
Chapter: The Cell: structure and function
Background
The Cell Theory
Scientific theories are well-substantiated explanations of the natural world acquired as a direct result of the scientific method, that have been extensively tested and confirmed. Biology has two: 1) the theory of evolution by natural selection and 2) the cell theory. The cell theory has three main components:
The cell is the basic unit of life
A cell (by definition) is bound by a cell membrane. This membrane provides a homeostatic environment within the cell, in which allows the internal environment to maintain near constant biochemical conditions. While much of the biochemistry that exists within a cell can be replicated outside of a cell, the presence of the cell membrane allows these chemical reactions to occur with high regularity and predictability. The cell is the basic unit of life indicates that it is the smallest unit considered to be alive. For example, DNA can be replicated outside of the cell, but DNA itself is not considered to be alive.
All living organisms are composed of cells
Organisms can be composed of one (unicellular) or many (multicellular) cells. This component of the cell theory simply indicates there are no other known units of life present in the known world. Viruses (which lack a membrane) are not be considered to be alive, even though they exhibit many (if not all) of the other properties of life, commonly accepted by biologists. Viruses have order, respond to their environment, and use energy to grow, develop, reproduce, and they evolve. Whether or not viruses are alive is debatable, but most biologists argue they are not, and invoke the cell theory to support this claim.
All cells came from other cells
Without an ability to reproduce, life would not exist. While the idea to life comes from life (known as biogenesis) seems obvious to us today, it was only widely accepted in 1864 after Louis Pasteur disproved the widely held belief (originally posited by Aristotle) that life emerges from non-living things, known as spontaneous generation (or abiogenesis). For example, it was thought that fleas emerged from dust, or maggots emerged from meat, spontaneously.
The Domains of Life
All cells have have few things in common. Cells are all bounded by a cell membrane, which consists of a phospholipid bilayer and embedded proteins. The cell membrane generates a regulated internal environment that allows the complex biochemistry for life to occur, within a gel-like fluid within the cell, known as a cytoplasm. All cells have DNA, which is a molecule that stores genetic information. The main functions of DNA are 1) to code for the production of proteins and 2) to replicate to allow cellular reproduction. According to the central dogma of molecular biology, DNA is the storage molecule that codes for the production of proteins. However, DNA does not code for proteins directly. Rather, DNA (a nucleic acid) produces a genetic message in RNA (a nucleic acid), which codes for the production of a protein at a ribosome. In other words, DNA synthesizes RNA, and RNA codes for the production of proteins. Therefore, all cells are bound by a cell membrane, which encases DNA which codes for the production of ribosomes (make up of RNA and proteins) and proteins. One fascinating discovery scientists have made is that all organisms on earth follow the same code. What does this mean? If you have an exact copy of a segment of DNA in different organisms, it will produce exactly the same protein. This is how genetic engineering is accomplished, a gene (a segment of DNA that codes for a specific mRNA molecule) from one organism can be inserted into a different organism, and once the gene is activate the genetically modified organism will produce exactly the same protein. The fact that all organism share the same code is a very strong indicator that all organisms on earth share a single, common ancestor. Another piece of evidence that supports this is that DNA replication is virtually identical for all organisms.

Figure 1. The three domains of life. All living things are composed of cells that contain DNA, which codes for proteins. Extant (currently living organisms) are placed into one of three domains. The domains Bacteria and Archaea, known as prokaryotes, lack a true nucleus and organelles. Whereas Eukarya, known as eukaryotes, have a true nucleus and true organelles. DNA evidence continually supports the hypothesis that eventhough, Archaea and Bacteria are more morphologically similar, Archaea is actually more genetically related to Eukarya. Bacteria differ from Archaea and Eukarya in the presence of a cell wall made up of the polysaccharide, peptidoglycan.
Traditionally, organisms were divided into two groups depending on the presence or absence of a nucleus: the prokaryotes and the eukaryotes. Prokaryotes (originating from the greek prokarya, meaning "before the seed" or "before the nucleus) form two of the three main branches of the tree of life: Bacteria and Archaea (Fig. 1). Prokaryotes are single-celled organisms that lack a true nucleus and membrane-encased organelles. Bacteria represent the oldest living organisms on the planet. Bacteria differ from Archaea in a number of ways, but morphologically the differ by the presence or absence of a cell wall. Bacteria have a cell wall made of a polysaccharide, called peptidoglycan. Archaea do not have this structure. Originally, it was thought that all prokaryotes were more closely related due to their similar morphologies. Furthermore, it was thought that Archaea preceded Bacteria, predominately due to the fact that it has a more simple structure. However, genetic analyses place Archaea more contemporary with the eukaryotes of Domain Eukarya (greek for "true seed" or "true nucleus"), which have a nucleus and complex organelles, than with their morphologically similar cousins, Bacteria. In fact, one of the only things that Archaea and Eukarya share in common, but differ from Bacteria, is the lack of a cell wall made of peptidoglycan. There are two solutions to explain this phylogeny. At some point in the distant past the common ancestor of all life diverged into two groups. One of those groups became Bacteria, while the other group eventually became Archaea and Eukarya. One hypothesis is that Bacteria (containing peptidoglycan) represent the oldest living clade, and the ancestor of Archaea and Eukarya had lost the cell membrane made of peptidoglycan. Alternatively, the ancestor of the original split between Bacteria and Archaea/Eukarya did not have peptidoglycan. Following the initial divergence, early bacteria evolutionarily developed a cell wall with peptidoglycan. Such a development could have been highly advantageous, allowing bacteria with the peptidoglycan cell wall to survive and reproduce, while those bacteria without peptidoglycan went extinct. These two hypotheses are highly contested.
Prokaryotes

Figure 2. Prokaryotic cell structure. While prokaryotes (i.e. bacteria) don't have organelles (which by definiition are bounded by a phospholipid bilayer), the do have several identifiable structures. Prokaryotic DNA is circular, and sometimes aggregates into a false nucleus, known as a nucleoid. It is not a true nucleus, as it lacks a nuclear membrane (a double phospholipid bilayer. DNA is transcribed producing mRNA which is immediately translated into proteins at the ribosome. All cells are bounded by a plasma membrane (or cell membrane). Bacterial cells have an additional cell wall beyond the plasma membrane. Bacterial cell walls are composed of the polysaccharide, peptidoglycan, which is only found in bacteria. Archaea is a separate group of prokaryotes from bacteria, which lack a cell wall.
Like all cells prokaryotes (Figure 2) are bounded by a cell membrane (or plasma membrane) containing the gel-like cytoplasm within, and have DNA that codes for the production of ribosomes and proteins. DNA in prokaryotes (Bacteria and Archaea) is not bound within a nucleus, while in eukaryotes DNA is bound within a nuclear envelope. In certain species of prokaryotes, the DNA is constrained within the cell, producing a structure which is visible under a microscope, known as a nucleoid. A nucleoid is not a true nucleus, in which DNA is constrained by a nuclear envelope. While prokaryotes have definitive structures they lack true organelles, defined by the presence of a phospholipid bilayer. Eukaryotes have true organelles. Bacteria do possess compartments of proteins not enclosed within a plasma membrane, thought to act similarly to primitive organelles.
Prokaryotes are typically unicellular or colonial
Prokaryotes are almost exclusively free-living, single-celled organisms. Some prokaryotes, such as cyanobacteria, can form large colonies. Such colonial bacteria have specialized proteins that allow the individual cells to connect with each. There are rare examples of bacteria that exhibit multicellular behavior during certain times of their life cycle, such as myxobacteria. When food sources are ample, these bacteria exist as unicellular organisms. As food supply decreases, myxobacteria swarm as colonies, as the individual cells attach to each other and move a singular unit. While the myxobacteria is swarming, they represent a colonial species, in which all the cells are similar in structure and function, yet connected. If the swarming colony of myxobacteria is unable to find a suitable food source, something very interesting happens. The swarming colony will halt its search and begin to form fruiting bodies. Within the fruiting bodies, some of the normal (vegetative, rod-shaped) cells change their structure into rounded, reproductive cells with extremely thick cell-walls, known as myxospores, which allows these cells to be more likely to survive until conditions become better. These reproductive cells are held atop the fruiting bodies by the carcasses of many of vegetative myxobacteria. At this point during its life cycle, myxobacteria exhibit very primitive multicellular behavior: the presence of different cell types. Certain eukaryotes, known as slime molds, have developed a very similar life cycle.
Protein synthesis in prokaryotes
The shape of DNA differs in prokaryotes and eukaryotes. In prokaryotes, DNA is a single, circular loop. In eukaryotes, DNA is organized into several linear chromosomes. In prokaryotes when proteins are required, a segment of DNA is transcribed into mRNA. As the mRNA is being transcribed, a ribosome attaches to the mRNA and begins the process of translation, converting the message of mRNA into a protein. Transcription and translation happen simultaneously in prokaryotes.
Eukaryotes
Eukaryotes are organisms made up of one or more cells that possess a membrane-bound nucleus (that houses DNA) as well as membrane-bound organelles. In contrast to prokaryotes, DNA is organized into linear chromosomes. Eukaryotic organisms may be multicellular or single-celled organisms. Unicellular eukaryotes are known as protists (which include amoeba, algae and dinoflagellates). Protists tend to be much larger (10-1000x) than prokaryotes, and like all eukaryotes have a true nucleus and organelles. Within Eukarya, multicellularity arose at least three times giving rise to modern day plants, animals, fungi.
Origin of the endomembrane hypothesis
Eukaryotes are thought to have emerged from an ancestral archaean, due to their close proximity suggested by Carl Woese's phylogeny (Fig. 1). While all prokaryotes have circular DNA, all eukaryotes have linear DNA. Therefore, the cell that is ancestral to all eukaryotes must have had linear DNA. At sometime in the past, the ancestral archaean, circular DNA was cut and made linear. This actually posed a significant issue in DNA replication, due to issues of chromosome shortening. This was resolved with the emergence of telomerase, of a protein/RNA hybrid, that allowed the ends of the chromosomes to be fully replicated.
The predominant nuclear origination hypothesis suggests infoldings of the cellular membrane (also called the plasma membrane) of an ancestral archaean (Fig. 3a). Imagine a group of brown paper bags. If you began to compress them together, the paper would begin to fold in on itself. This is a good metaphor for the infolding hypothesis. Evidence to supports this comes from the presence of such infoldings in prokaryotic species that multiply in constrained areas (Fig. 3b). Unlike a paper bag, the cellular membrane is composed of phospholipids, which are capable of detaching from the outer membrane. The nuclear origin hypothesis suggests that the detached infoldings condensed and encased the DNA as the phospholipids reattached to each other (Figure 3), forming nuclear membrane and the endoplasmic reticulum, which form a contiguous system, with the these organelles physically connected to each other. The nucleus and the endoplasmic reticulum are a part of the endomembrane system, within eukaryotic cells, which also include the Golgi apparatus, vesicles and lysosomes (Fig. 4). These organelles are thought to have emerged in a similar fashion (possibly synonymously) as the nucleus and endoplasmic reticulum, but are separate from the nucleus and endoplasmic reticulum.

Figure 3. Origin of the endomembrane hypothesis. a) Eukaryotes emerged from an ancestral archaean cell. DNA is some prokaryotes are concentrated within the cell, in an area known as a nucleoid. b) The cell membrane of the ancestral archaean cell began to infold on itself forming plasma membrane within the cell. We see this in certain species of bacteria, and in some cases these infolding isolate the nucleiod.. c) The first true nucleus formed with the infoldings separated from the cell membrane, and encased the chromosomes, creating a nuclear envelope. The endoplasmic reticulum is physically connected to the nuclear envelope, yet reaches out into the cytoplasm of the cell. Additional organelles of the endomembrane system include the Golgi apparatus, vesicles and lysosomes.
The nucleus
.svg)
Figure 4. Endomembrane system of a eukaryotic cell. Linear DNA, known as chromosomes, housed within the nucleus generates RNA molecules. Processed RNA molecules exit the nuclear envelope via nuclear pores and enter into the rough endoplasmic reticulum (or the cytoplasm) and attach to a ribosome (synthesized in the nucleolus of the nucleus), where a protein is synthesized. Attached to the rough endoplasmic reticulum is the smooth reticulum, which lacks ribosomes and is responsible for lipid synthesis. Once a protein is synthesized, it may travel to the Golgi apparatus along the cytoskeleton network. At the Golgi apparatus, the protein can be modified and packaged for use within the cell or excreted. Lysosomes are specialized vesicles that are responsible for breaking down a variety of biochemicals.
Unlike prokaryotes, eukaryotes have a true nucleus (Figure 4), defined as DNA housed inside of a nuclear envelope, also called the nuclear membrane. The nuclear envelope is a double-membrane composed predominately of phospholipid bilayers, and is punctuated by nuclear pores. These openings in the nuclear envelope allow the movement of specific molecules into and out of the nucleus. Namely, mRNA leaves the nucleus through the nuclear pores and nucleotides enter into the nucleus in order to create new DNA and RNA. Inside the nucleus several strands of linear DNA, known as chromosomes, are housed. When you look at the nucleus under the microscope, you will see a dark spot within the nucleus, the nucleolus, which is the ribosome-manufacturing facility.
It is thought that the primary advantage the nucleus provides is gene regulation, by separating transcription and translation. Transcription is the process in which a segment of DNA (known as a gene) codes for a messenger RNA (mRNA). Following transcription, eukaryotes undergo post-transcriptional modification in which segments (introns) of the unprocessed mRNA (or primary transcript) are removed prior to leaving the nucleus, and the remaining segments (exons) are reattached, forming a mature mRNA. The mature mRNA leaves the nucleus through a nuclear pore and travel to a ribosome. The ribosome reads the mature mRNA and with the help of transfer RNA (tRNA) links amino acids in a specific sequence generating a protein. Prokaryotes (which don’t have a nucleus) go through transcription and translation simultaneously, and therefore are incapable of gene regulation via post-transcriptional modification.
Endoplasmic reticulum
The endoplasmic reticulum is an endmembrane organelle contiguous with the nuclear envelope that consists of a series of flattened, interconnected membranes. Rough endoplasmic reticulum houses ribosomes and is an important location for the synthesis of proteins, vesicles and lysosomes. Smooth endoplasmic reticulum lacks ribosomes, and therefore is incapable of protein synthesis, but primarily functions as the production center for lipids, namely: phospholipids, lipids and steroids.
In review, within the nucleus DNA synthesizes RNA molecules via transcription, in which a precise segment of DNA (known as a gene) opens up and synthesizes a specific strand of mRNA (refer to Figure 6). After the mRNA is synthesized, post-transcriptional modification occurs in which segments (introns) of the mRNA and the remaining mRNA segments (exons) are reattached. This mature mRNA (composed of reattached exons) leaves the nucleus through the nuclear pore and either enters into the cytoplasm or the rough endoplasmic reticulum (rough ER). Either in the cytoplasm or the rough ER, the mature mRNA attaches to a ribosome. At the ribosome, the process of translation begins in which the mRNA codes for the synthesis of a protein. Once the protein is completely synthesized at the ribosome, the protein leaves the rough ER through a transport vesicle.
Vesicles
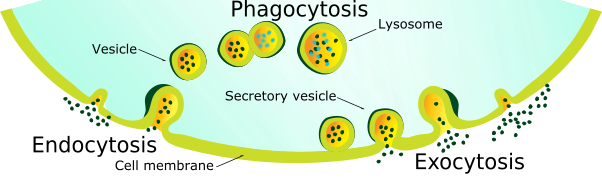
Figure 5. Endocytosis, pinocytosis and exocytosis. In endocytosis, larger particles can enter the cell in pockets of the cell membrane. These pockets pinch together forming vesicles. Lysosomes fuse with these vesicles and break down smal biological molecules (pinocytosis) or food particles (phagocytosis). Lysosomes can also fuse with the cell's own defective organelles and recycle in their material for reuse, in a process known as autophagy. Materials destined for excretion are packaged in secretory vesicles, which fuse with the cell membrane, releasing the waste material into the intermembrane space.
Vesicles are organelles within a cell, consisting of fluid enclosed by a phospholipid bilayer. Vesicles perform many different functions within the cell: the movement of substances into and out of a cell, as well as movement of substances within a cell, and as storage units. Certain large biomolecules, such as proteins are incapable of passing across the cell membrane via passive transport mechanisms, such as diffusion. In endocytosis, the cell membrane engulfs larger molecules via active transport (Figure 5). These molecules collect in pockets of the phospholipid bilayer, which grow inward, using energy (ATP). As the pocket of the cell membrane grows inward, the phospholipid bilayer begins to pinch together toward the exterior of the cell. Eventually, the pocket detaches from the cell membrane and becomes a vesicle. Since vesicles are made up phospholipids, they are capable of merging with (and emerging from) other membranes of organelles, as well as the cell membrane. In exocytosis, a form of active transport, a secretory vesicle fuses with the cell membrane releasing waste material (Figure 5). Transport vesicles move biological molecules from one organelle to another, using energy (ATP). For example, certain proteins are transported within a transport vesicle from the rough endoplasmic reticulum to the Golgi apparatus for further processing.
Vacuoles
Vesicles do more than move substances into, within and out of the cell. Since vesicles are separated from the cytosol, the main body of fluid within the cell, vesicles can have a different biochemical environment. Vacuoles are specialized vesicles that hold predominately water and can be found in many eukaryotes, including plants and animals. In animal cells vacuoles are very small, while in plant cells the vacuole is typically the largest organelle of the cell. In most plant cells, the vacuole allows the cell to maintain osmotic balance and nutrient storage. Vacuoles also store pigments in the cells of colorful plant organs, such as flower petals, providing them color. Vacuoles within plant seeds can store proteins and fats, which serve as a rich energy source for developing seeds.
Lysosomes

Figure 6. Protein synthesis, modification and shipment. DNA codes for the production of mRNA within the nucleus. The mRNA exits the nuclear envelope through a nuclear pore and enters into the rough endoplasmic reticulum, where it attaches to a ribosome and a protein is synthesized. The protein leaves the rough ER through a transport vesicle and enters the cis-face of the Golgi apparatus, where it is modified and identified within the cisternae of the Golgi. The modified protein exits the Golgi through the trans-face via a secretory vesicle, where (depending on the protein) it either attaches to another organelle or cell membrane, or it gets excreted from the cell via exocytosis.
Lysosomes are specialized vesicles involved in cellular digestion. They contain a variety of enzymes that allow them to break down a wide array of biological molecules engulfed by the cell. In a process known as phagocytosis (Figure 5), vesicles containing food enter into the cell via endocytosis fuse with lysosomes, which release their digestive enzymes into the merged organelle (Figure 5), breaking down biomolecules such as carbohydrates, lipids, nucleic acids and proteins into their fundamental monomers. These monomers can be used by the cell's mechanisms to formulate its own unique biomolecules (i.e. DNA and proteins). In addition to digesting food particles, lysosomes assist the cell by recycling its own defective organelles and biomolecules, in a process known as autophagy. Once these structures are broken down, the cell can reassemble the monomers to form new biomolecules and organelles. Lysosomes are also thought to be involved in pre-programmed cell death in multicellular species. As organisms develop from a single cell into an adult, they take on many different forms. It is thought that this change occurs by the predetermined death of certain cells during specific times of development. For example, at a very early stage of human development, the embryos have tails and webbed hands and feet. As the fetus develops, the tail begins to shrink and the webbing between the fingers and toes disappears. This happens because those cells are killed by their very own lysosomes, but aids in the overall development of that organism.
Golgi apparatus
Some synthesized proteins require further processing and are transported to the Golgi apparatus to be modified. A transport vesicle from the rough endoplasmic reticulum with the unmodified protein travels to the cis-face of the Golgi apparatus along the cell’s cytoskeleton network. The Golgi apparatus is a collection of flattened membranes known as cisternae. Once the transport vesicle containing proteins enters into the Golgi apparatus, some of the the peptide bonds are broken and rearranged, creating an altered protein. In addition, identification tags are put on the protein that allow it to be placed exactly where it is designed to be. The modified protein leaves the Golgi apparatus through the trans-face and connects to the cytoskeleton network, traveling to its identified location. Proteins packaged in transport vesicles are either destined for secretion via exocytosis or to be used by the cell. The Golgi apparatus can be thought of as a post office, in which proteins are labeled, packaged and shipped to the appropriate destination.

Figure 7. Cell movement in unicellular organism by cilia and flagella. One function of the cytoskeleton allows unicellular organisms to move. Cilia are multiple, small protuberances of filaments that emerge beyond the cell membrane, which beat in a wave-like fashion allowing cell movement and move nutrient-rich water across the surface. Larger filaments, known as flagella, extending from the cytoskeleton whip in a back and forth motion allowing for a highly efficient form of mobility.
Cytoskeleton network
The cytoskeleton is an interconnected network of protein filaments that exists in all cells, including prokaryotes and eukaryotes. The cytoskeleton serves many different functions within the cell. Its primary function is to provide resistance against compression allowing the cell to maintain its overall shape (akin to an animal's skeleton). This allows unicellular growing next to each other resistance to mechanical deformation. In multicellular organisms, this resistance allows tissue stabilization and structural integrity.
The cytoskeleton is also involved in cell movement. The proteins of the cytoskeleton are capable of contracting and releasing, which alter the shape of the cell allowing the cell to move. For example, in muscle cells actin filaments contract to shorten the cell. In some cells, the cytoskeleton extends beyond the cell membrane, and either form cilia or flagella (Figure 7). Cilia are protuberances that project beyond the body of the cell, and are common in many different types of cells. Certain prokaryotic and eukaryotic unicellular species have cilia that beat in coordinated waves allowing them to maneuver through water or to move water over the cell surface as a feeding mechanism. This is an example of motile cilia. Motile cilia also exist in multicellular species. For example, cell inside the trachea of vertebrates beat continuously to remove mucus and dirt particles out of the body. Motile cilia in the Fallopian tubes in humans are responsible for carrying the egg from the ovary to the uterus. Flagella are typically much longer, thicker but less numerous than cilia. These extreme extension of the cytoskeleton whip back and forth creating an effiencent locomotion mechanism.
In eukaryotes, the cytoskeleton network is also responsible for the movement of vesicles within the cell. Once a protein is created by a ribosome in the rough endoplasmic reticulum, it leaves as the ER pinches off, forming a transport vesicle. This transport vesicle moves along the cytoskeleton filaments towards the Golgi apparatus. Once processed, the protein leaves the Golgi apparatus in a vesicle to be transported with or out of the cell via exocytosis. The cytoskeleton also plays a role in endocytosis and organelle transport.
The cytoskeleton is critical in cell division. Prokaryotes divide in a process known as binary fission. Following the replication of DNA, the filaments of the cytoskeleton constrict migrating the DNA strands to opposite poles of the cell. Another filament constricts the middle of the cell, pinching together the cell membrane and cell wall, eventually producing two cells. During mitosis and meiosis in eukaryotes, filaments emerge from centrioles and attach to the chromosomes. These filaments constrict, eventually separating the paired chromosomes and pulling them to opposite poles. Once this occurs, a different filament constricts the cytoplasm, in a process known as cytokinesis, dividing the mother cell into two daughter cells.

Figure 8. Structures of a mitochondrion. The mitochondrion is a double-membraned organelle within eukaryotes involved in aerobic respiration. The outer membrane have proteins, called porins, that allow up to medium sized molecules in and out of the mitochondrion, generating an aqueous solution in the intermembrane space similar to the cytosol. However, large proteins made by the mitochondrion remain. The inner membrane is highly folded into structures known as cristae and is primarily responsible for oxidative phosphorylation in the electron transport chain, with the protein, ATP synthase, ultimately generating most of the ATP produced during cellular respiration. The environment inside the inner membrane, known as the matrix, contains a variety of enzymes (most notably those responsible for the citric acid cycle) and residual mitochondrial DNA capable of synthesizing its own RNA and proteins.
Mitochondrion
The mitochondrion is the organelle responsible for cellular respiration in nearly all eukaryotes. Mitochondria are organelles that have been described as the powerhouse of the cell because they synthesize most of eukaryotes' Adenosine Triphosphate (ATP), the chemical source of energy used by all organisms. Mitochondria are composed of several compartments (Figure 8) that serve specific functions. Mitochondria have two membranes (an inner and outer membrane) and an intermembrane space between. While the outer membrane is spherical to oblong shaped, the inner membrane is folded in on itself forming several invaginations, known as cristae. The outer membrane has protein complexes known as porins, that allow small to medium sized molecules to freely diffuse into and out of the organelle. Therefore the intermembrane space is quite similar to the cytosol, with the exception of large molecules (specifically proteins) that are incapable of moving across the outer membrane. The foldings, or cristae, of the inner membrane increase the surface area, which is important as this is where much of the biochemistry of the electron transport chain of cellular respiration occurs, including: the redox reactions of oxidative phosphorylation and the synthesis of ATP. The space inside of the inner membrane, known as the matrix, is where many biochemical reactions occur, including the citric acid cycle. Also housed within the matrix is DNA and ribosomes. But how?
Ancestral eukaryotes generated ATP by performing glycolysis within cytosol within the cell. All present day eukaryotes also go through glycolysis. However, the net production from glycolysis is 2 ATP. Eukaryotes with a mitochondrion can net up to 36 ATP, an energy efficiency of 16 times greater.
There are two hypotheses about the origin of mitochondria: endosymbiotic and autogenous. The endosymbiotic (when one organism likes inside another mutually benefiting both) hypothesis (Figure 9) suggests the mitochondrion was originally a prokaryotic cell engulfed by an early eukaryotic cell over 2 billion years ago. Whereas the autogenous hypothesis suggests that DNA from an ancestral eukaryote split and exited the nucleus, which was entrapped by a phospholipid bilayer, further engulfed by an additional bilayer, giving you two membranes. This must have occurred prior to the original divergence of the Bacteria from Archaea/Eukarya, and is retained as a remnant in eukaryotes. However, mitochondria have many similarities with bacteria causing most scientists to discredit the autogenous hypothesis in support of the endosymbiotic hypothesis.
Figure 9. Origin of the mitochondrion. According to the endosymbiosis hypothesis, an ancestral eukaryote (a) engulfed, but did not digest, a protobacterium (b). In an increasingly oxygen rich environment, this bacterium was capable of extracting much more energy (ATP) from biological molecules than the eukaryote alone. The protobacteium benefited from a supply of undigested food particles, and eventually became an endosymbiotic mitochondrion (c), living inside the eukaryote.
The endosymbiotic hypothesis posits a single unicellular eukaryotic cell (see Figure 3c) engulfed a bacterium (now known as the mitochondrion), approximately between 1.7 and 2.2 billion years ago. It is unclear whether this engulfment of the mitochondrion occurred before, during or after the origin of the nucleus. Instead of digesting the bacterium for food, the eukaryotic cell retained it. This proved to be one of the most important interactions for the evolution of life on Earth. Why? This eukaryotic cell could take one molecule of glucose and net 32 ATP, rather than 2 ATP prior to the engulfment. It is thought that the mitochondrion also benefited from this relationship in two ways. First, it got protection. Being housed in a much larger cell (100-1000x larger than itself), it was no longer preyed upon by bacteria-eating microorganisms. Second, the the eukaryote alone is incapable of digesting many biological molecules. This first mitochondrion flourished inside its host due to an abundance of half-eaten molecules. Perhaps the best evidence that the bacteria benefited from this relationship is that bacteria resembling the mitochondrion are extinct, but mitochondria are not. What is extraordinary to ponder is that this happened once….in a single cell. Following the emergence of the first ancestral eukaryote, this new organism was so successful that it multiplied and diversified at such a tremendous rate that scientists are still trying to understand how eukaryotes are related just beyond the root of Eukarya. Whatever happened, this single cell was so successful that it eventually gave rise to all protists, plants, fungi and animals on earth.
A significant body of evidence exists to support the endosymbiotic hypothesis. 1) Mitochondria are the same size as the average bacterium. 2) The strongest evidence is that mitochondria have their own DNA, and can manufacture some of their own, RNA, ribosomes and proteins. And in fact, if you sequence the DNA of the mitochondrion and compare it with taxa from the entire tree of life, the DNA sequence is most closely related to a bacteria (not a eukaryote). In addition, mitochondrial DNA is composed of multiple copies of the same, circular chromosomes. Eukaryotes, in contrast, have DNA that is organized as many different, linear chromosomes. 3) Mitochondria are double-membraned, most organelles are single-membraned. It is thought that when the original eukaryote engulfed the mitochondrion, the membrane of the original eukaryote (phagocytic vacuole) wrapped up the mitochondrion, like a bag within bag). 4) Interestingly, the internal membrane of the mitochondrion has a lipid composition more similar to bacteria, whereas the external membrane has a lipid composition more similar to eukaryotes. Only one other eukaryotic organelle has two membranes, and it is thought that this organelle was consumed by a descendent of this first aerobic eukaryote. That organelle was the chloroplast, which helped give rise to photosynthetic eukaryotes and eventually plants.

Figure 10. Comparison of a chloroplast and cyanobacterium. Chloroplasts are thought to have originated from a singular, endosymbiotic event in which a eukaryote engulfed, but did not digest, a cyanobacterium. In both structures, chlorophyll (the site of the light reactions) is housed on membranes of internal structures known as thylakoids. Membranes of thylakoids in cyanobacteria run parallel to the cell membrane. Whereas thylakoids in chloroplasts stack generating structures called grana, enhancing the internal surface area allowing for more chlorophyll and thus, greater efficiency. Both cyanobacteria and chloroplast have nucleoids, containing circular DNA capable of producing RNA and proteins. They also both have two membranes, likely a remnant of an endosymbiotic event. The fluid between the thylakoids and inner membrane, known as the stroma, is the location of the Calvin cycle.
Chloroplast
Chloroplasts are photosynthetic organelles found in plant and algal cells. The photosynthetic pigment, chlorophyll, captures light energy and converts it into chemical energy. Chlorophyll is housed on the membrane of pancake-shaped structures, known as thylakoids, within the body of the chloroplast. Thylakoids tend to stack on top of each other forming stacks, known as grana. Adjacent grana can be connected together by membranous bridges, called lumen. Light reactions of photosynthesis occur on the membrane of the thylakoid, splitting water (H2O). Energy is generated during this anabolic reaction, which is used to synthesize ATP. Single oxygen atoms bind to form oxygen gas (O2), a waste product. Hydrogens are captured by NADP to form NADPH. ATP and NADPH enter into the stroma, or fluid between the thylakoids and cell membrane. In the stroma, ATP is used to drive the Calvin cycle, in a series of catabolic reactions occurs where hydrogens are stripped from NADPH and rearranged with the atoms of carbon dioxide (CO2) to form the sugar, glucose (C6H12O6).
Approximately a billion years ago, a cyanobacterium entered into eukayrotic cell. It either entered as an internal parasite, or was engulfed similar to how the mitochondrion. However the cyanobacterium entered the cell, modern day chloroplasts do not have a eukaryotic phagocytic vacuole, which was either lost, in the case of engulfment, or never occurred, in the case of parasitism. Cyanobacteria (and primitive chloroplasts, i.e. glaucophytes) represent a type of bacteria known as Gram-negative, which have a double membrane with a cell wall sandwiched between, made of peptidoglycan, a polysaccharide absent in eukaryotes but present in bacteria (but not archaea). Thus it is posited the double membrane of the chloroplast represent the inner and outer membranes of the original cyanobacterium. However unlike the mitochondrion, there is no presence of a eukaryotic membrane, in contemporary plant cells.
Figure 11. Origin of the chloroplast. A heterotrophic, mitochondrion-containing eukaryote (a) was either parasitized by or engulfed a cyanobacterium (b), producing an autotrophic, photosynthetic eukaryotic cell (c).
This new structure gave the eukaryote the significant advantage of becoming autotrophic, being able to generate chemical energy from sunlight via photosynthesis. Prior to this eukaryotes were heterotrophic, in which they had to extract food from their environment either through filter feeding or predation. The eukaryotic host must have had mitochondria prior to the engulfment of the cyanobacterium, as all photosynthetic eukaryotes presently have mitochondria. Over time, most of the cyanobacterial DNA was either lost or assimilated into the nuclear chromosomes of the eukaryotic host. Most proteins needed by modern day chloroplasts are synthesized by the eukaryotic transcription and translation, and imported to the chloroplast. Some bacterial DNA (plastid DNA) persists within modern-day chloroplasts. At approximately 100 genes, the plastid DNA is capable of producing some of its own proteins, similar to (but far fewer than) the mitochondrion.
Most scientists concur that all chloroplasts within eukaryotes have been traced back to a singular endosymbiotic event, with one exception, even though many eukaryotes with chloroplasts are distantly related to each other. This suggests that the chloroplast endosymbiosis event occurred early in eukaryote evolution, but has been being lost several times over throughout evolutionary history. There are three chloroplast lineages (most primitive to most recent): glaucophyte (blue-green algae), rhodophyte (red algae) and chloroplastidan (green algae and land plants).
Plant cells vs. animal cells
Plants and animals are eukaryotic organisms. So they share a similar internal structure of organelles and other cellular structures (Figure 12-13). Plant cells differ in three main ways from animal cells. (1) Plant cells have photosynthetic organelles, chloroplasts, that appear green under a compound microscope. (2) While animal cells have vacuoles, a plant cell’s vacuoles are very large in comparison. The main function of plant vacuoles is to store water between precipitation events. (3) Plant cells have thick primary cell walls made of the polysaccharide, cellulose. Plant cells involved in structural support also have a secondary cell wall made of a very dense polysaccharide, lignin. This gives plants a roughly polygonal shape, whereas animal cells (which lack a cell wall) have a more amorphous shape.

Figure 12. A typical animal cell. Plant cells have many of same internal cellular structures and organelles as animals cells, with a few exceptions. While both animal and plant contain vacuoles useful for water and nutrient storage, a plant cell's vacuoles is enormous by comparison. This is a result of an immotile life style, and being dependent on rain. Animals in contrast are motile, and either live in water or are capable of moving to search for water.

Figure 13. A typical plant cell. In addition to large vacuoles, plant cells contain chloroplasts, whereas animal cells do not. Plant cells also have a cell wall made of the polysaccharide, cellulose that helps the cell maintain its rigid structure as plants grow.