
Plant Reproduction
Plant reproduction is the production of plant offspring, which happens either sexually or asexually. Asexual reproduction produces a genetic clone of the original plant, whereas sexual reproduction occurs by the fusion of gametes, producing offspring that are genetically unique from the parent plants. Sexual reproduction recombines the alleles from one generation to the next, enhancing the population’s genetic diversity. Increasing a population’s genetic diversity via sexual reproduction allows the population of plants, as a whole, to adapt to varying environmental conditions, be they biotic (i.e. diseases) or abiotic (i.e. a localized disturbance such as drought or longer term impacts such as climate change). However, sexual reproduction requires a successful transmission of gametes from one individual plant to another, which is challenging considering the immobility of plants. To minimize the risks of sexual reproduction, many plants are capable of reproducing sexually or asexually. Asexual reproduction allows plants to reproduce in the absence of fertilization, which allows them to expedite localized expansion. The major risk of asexual reproduction is newly formed plants produced are genetically identical (barring mutations) to the parent. While this may be an advantage in the short term, environmental conditions are constantly changing and species with low genetic diversity due to a heavy reliance on asexual reproduction may dies in large numbers if the environment suddenly changes or a new disease enters into the population.

Asexual Reproduction
Asexual Reproduction
Asexual reproduction produces new individual plants that a genetically identical to the parent plant, unless mutations have occurred in the originating cells. Asexual reproduction can happen in plants by two different mechanisms: vegetative reproduction and apomoxis. Vegetative reproduction is an example of asexual reproduction, in which a vegetative portion of the plant (i.e. leaf, stem or root) gets removed from the parental plant and generates a separate individual. Apomoxis occurs when unfertilized gametes (pollen or ovules) develop into genetically identical spores or seeds.
Types of Vegetative Reproduction
Vegetative reproduction happens by a variety of mechanisms in plants. Many plants have modified stems that specialize in the development of new individual plants, that once established are capable of living away from the parent plant. Rhizomes are modified stems that run underground and emerge away from the parent plant producing a new individual. Iris plants commonly reproduce this way forming colonies (or clumps). Aspen trees (within the genus Populus) grow in extremely large colonies, which predominantly reproduce via rhizomes which typically travel below ground for 30-40m from the parent plant. One colony of aspens in Utah is estimated to be over 80,000 years old, and considered to be the largest organisms in the world. Plants that reproduce via rhizomes are also resistant to fire. Temperatures just below the soil line are highly mediated during a fire event, and these plants typically are capable of resprouting new plants after a fire. Stolons are another example of modified stems that promote vegetative reproduction. Rather than spreading below ground, stolons (commonly known as “runners”) spread away from the parent plant above ground. The most common example of this is crab grass used in lawns. Strawberries also reproduce via stolons. Plants can also vegetatively reproduce from their root system. Roots and stems are anatomically different, so a rhizome is distinct from a root. In certain species (i.e. roses), as the root system reaches the surface of the soil profile, adventitious buds emerge forming stems and leaves, and subsequently a new plant. This process is known as suckering. Certain species that produces underground bulbs (i.e. onion and tulips) can vegetatively reproduce by producing new bulbs. Bulbs are highly specialized underground plant structures that consist of a short stem with underground leaves. Typically, a bulb is a storage mechanism that allows the plant to lie dormant during the winter season. Once spring emerges the underground leaves emerge and begin to photosynthesize.
Types of Apomixis
Apomixis is the development of unfertilized embryo into a new individual plant. There are many variations of apomixis among plants. Nearly all plants that undergo apomixis are facultative (facultative apomixis), meaning that apomixis only occurs in gametes that are not successfully fertilized. In certain species of non-vascular plants (i.e. moss, liverworts and hornworts) and a few seedless vascular plants (i.e. ferns and club mosses), what appear to be gametophytes (structures that produce haploid gametes) actually produce cells that look like sporophytes (haploid cells: egg or sperm) but retain the same chromosome number (ploidy level) as the parent plant. This process of apomixis is known as apogamy, and produces genetically identical spores (single-celled embryos).As far as we know, apomixis is completely absent in gymnosperms. However, there are several variations of apomixis in angiosperms (flowering plants). In gametophyic apomixis, a diploid embryo develops into a seed (a multicellular structure including the embryo and stored food in the endosperm) from a cell that did not undergo meiosis. Some species of onion (Allium sp.) undergo vegetative apomixis, in which the unfertilized embryos of the flowers develop into vegetative structures (known as bulbils) on the inflorescence. These bulbils resemble small plants, with stems, roots and leaves present. Once the bulbils contact moist soil, they can develop into a new individual.
Sexual Reproduction
Sexual reproduction in plants results from fertilization, the union, of gametes (cells that have undergone meiosis) from two genetically different plants. Meiosis reduces the number of chromosomes in half producing gametes. Fertilization recombines gametes from different individuals, producing a zygote, which develops into either spores or seeds. The main purpose of sexual reproduction is the rearrangement of genes in the next generation. This serves to increase the genetic diversity of the population, enhancing the evolutionary viability of the species in the face of changing environmental factors.
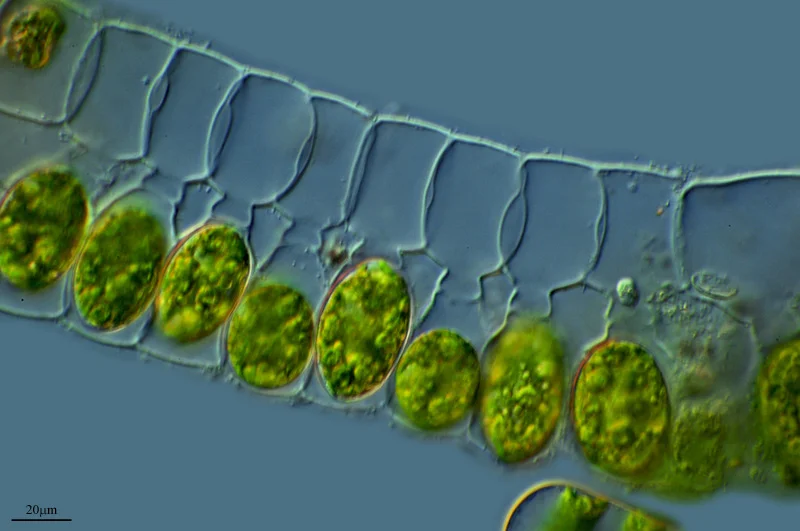
Conjugation in green algae
Conjugation: the origin of sexual reproduction in green algae
Green algae (Phylum Chlorophyta) are aquatic, photosynthetic species that can be unicellular, colonial (cells of all the same type attached to one another) or multicellular (different types of cell types within an organism). While most green algae reproduce via asexual reproduction, certain groups (notably Spirogyra) developed sexual reproduction. In a process known as conjugation, a diploid (with 2n chromosomes) green algal cell will undergo meiosis producing four haploid (n chromosomes) cells that recombine via fertilization to form genetically unique diploid zygotes in the next generation. During conjugation, a fertilized diploid (2n) algal cell begins this process by thickening its cell wall producing a diploid zygospore with a spiny outer shell. The zygospore then undergoes meiosis producing four haploid (n) cells inside the spiny cell wall. Two of these haploid cells will be of one mating strain (+), while the other two are of the opposite mating strain (-). During fertilization, + mating strains can only fuse with – mating strains. This is a primitive version of female and male gametes later developed in plants. Once the haploid cells have developed, the spiny cell wall of the breaks apart in a process known as germination, freeing the haploid cells. Following germination, the haploid cells swim with two flagella and aggregate together forming clumps. Each clump includes haploid cells of the same mating type, generating + mating type clumps and – mating type clumps. When the different mating type clumps come in contact with each other, a cellular membrane tube (known as a cytoplasmic thread) forms between a + haploid and a – haploid cell. As the cytoplasmic thread enlarges, the cytoplasm and haploid (n) nuclei of the two cells merge forming a genetically unique diploid (2n) green algal cell, in a process known as fertilization.

Alternation of generations
Alternation of Generations originated in multicellular green algae
Whereas conjugation is a form of sexual reproduction within a unicellular or colonial species, certain groups of multicellular green algae (and land plants) reproduce by a process known as alternation of generations. In this process, there are two multicellular generations linked by a unicellular reproductive stage. One generation, known as a gametophyte, produces an entire haploid (n) multicellular organism and produces haploid (n) gametes via mitosis in a specialized tissue known as a gametangium, a gametophyte producing gametes with exactly the same number of chromosomes as the somatic (or body) cells of the organism. Haploid (n) gametes from the gametophyte generation fuse to generate a diploid (2n) zygote. As the zygote develops into a diploid (2n), multicellular organism, known as a sporopyhte. Unlike the gametophyte stage, the sporophyte produces spores by undergoing meiosis, reducing the number of chromosomes in half, in a specialized tissue known as a sporangium. These haploid spores germinate producing the haploid gametophyte, starting the cycle again. Multicellular green algae are neither gametophyte dapoominant, nor sporophyte dominant. Land plants are either gametophyte dominant or sporophyte dominant.
Stoneworts have multicellular reproductive structures
Stoneworts (Class Charophyceae) are aquatic photosynthetic organisms, which represent a developmental step between green algae and land plants. Stoneworts have vegetative tissues that include whorls of filaments centered on a node connected by a cylindrical axis, and contain single-cell, root-like structures (rhizoids) that allow them to attach to a substrate. Unlike multicellular green algae, extant stoneworts do not appear to reproduce via alternation of generations. Reproductive organs attached at the axial nodes include the oogonium and antheridium. Oogonina and antheridia are both surrounded by an envelope of sterile (infertile) cells. The oogonium is a multicellular, female sex organ that contains one, relatively large immotile egg, known as a megaspore. The oogonium is a large vase-shaped (or oval) structure made up an envelope of spiraling filaments, whereas the antheridium is a smaller globular structure. Separate male sex organs (antheridia) each contain one relatively small sperm cell, known as a microspore. Gametes are protected within a multicellular structure are not seen in green algae, which is the main argument against their inclusion in Phylum Chlorophyta. Relative to green algae, stoneworts have extremely large eggs. Motile, sperm cells (n) swim to immobile egg cells (n) fertilizing them creating a diploid (2n) zygote, which can germinate producing a new genetic, diploid individual. Species within Charophyceae are either monoecious (antheridia and oogonia exist on the same plant) or dioecious (individuals either produce antheridia or oogonia, but not both).
Embryos of land plants are enveloped by nutritive, protective tissues
True plants (Class Embryophyceae) have several morphologies that allow them to persist on land. Reproductively, land plants are distinct from stoneworts in that plant embryos extract nutrition from the tissue surrounding them, enhancing their ability to survive during their early stages of development. In addition to nutritive tissue, plant embryos have a protective outer tissue that help the embryo from dehydrating while on land. While extant (still living) stoneworts do not appear to reproduce via alternation of generations, all land plants do. Most taxonomists conclude land plants emerged from stoneworts (due to genetic analyses and the multicellular nature of the reproductive organs), suggesting that ancestral stoneworts likely reproduced via alternation of generations, but those taxa are now extinct.
Bryophytes transitioned from water to land
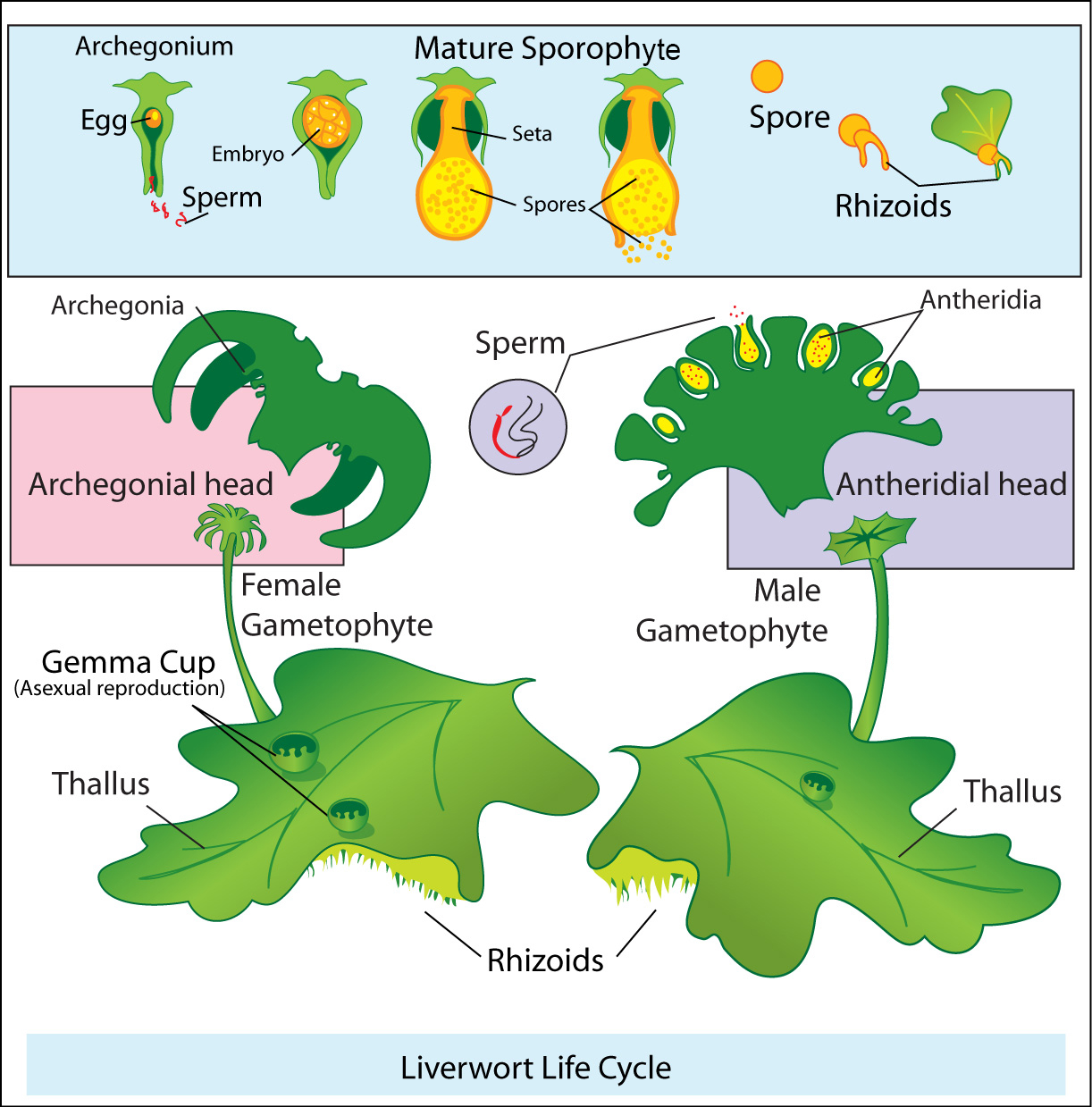
Bryophytes (or non-vascular plants) emerged from stoneworts and represent the first true land plants. Bryophytes are small land plants lacking true vascular tissue, reproduce via spores, and include true mosses (Phylum Bryophyta), liverworts (Phylum Hepatophyta) and hornworts (Phylum Anthocerotophyta). Transitioning to land required unique morphologies not found in stoneworts allowing plants to retain an adequate osmotic (H2O) balance. Vegetatively, land plants have a waxy cuticle layer on the exterior of leaves (and certain stems, i.e. cacti), which creates an air-tight system preventing water from diffusing out of the plant. However, plants need CO2 to undergo photosynthesis. Specialized cells, known as guard cells, exist in the epidermis of the leaf that open and closes, creating pores in the leaf known as stomata. While this allows CO2 in, it also releases water out. Plants are constantly opening and closing their stomata to regulate osmotic balance inside the leaf depending on current soil moisture availability. All land plants reproduce via alternation of generations. Haploid gametophytes have cells with several unpaired chromosomes and produce haploid gametes via mitosis. These gametes fuse during fertilization, producing a zygote that develops into a sporophyte, containing diploid somatic cells. Specialized tissues undergo meiosis, producing four haloid spores, which germinate without fertilization developing into a gametophyte.
The bryophyte life cycle is gametophyte dominant, meaning that the majority of the life cycle is dominated by the gametophyte generation. Gametophytes are more prominent and longer-lived than the sporophyte generation. Sporophytes emerge intermittently, and are typically nutritionally dependent upon the gametophyte for development. Gametophytes of bryophytes are either male or female. While the photosynthetic tissues of gametophytes are virtually identical, the reproductive organs are unique to each sex and housed either on the tips of the shoots or in the axils of the leaves. Male gametophytes produce sexual organs, known as antheridia, which are ovular structures, made of several male gametes, or sperm encased by a sterile, infertile tissue. In contrast, a stonewort’s antheridium produces only one male gamete. Female gametophytes produce a vase-shaped sex organ, known as an archegonium, with a single egg house inside a sterile tissue. The male gametes of bryophytes (as well as green algae and stoneworts) require water to reach the archegonium. In the presence of water, male gametes of bryophytes swim toward the antheridium guided by a sexual pheromone (chemical) produced by the archegonium.
Once the male and female gametes fuse via fertilization, they produce a diploid zygote, known as a sporophyte embryo. This embryo develops inside of the archegonium, eventually producing a sporophyte. Sporophytes are typically inconspicuous and non-photosynthetic. As a consequence, they are nutritionally reliant on the gametophyte and remain attached. At the end of the sporophyte development a multicellular structure develops, known as a sporangium, in which the internal cells undergo meiosis producing male and female haploid (n) spores. These spores are typically wind dispersed and germinate developing into a gametophyte restarting the bryophyte life cycle.
Megagametophytes and microgametophytes in lycophytes
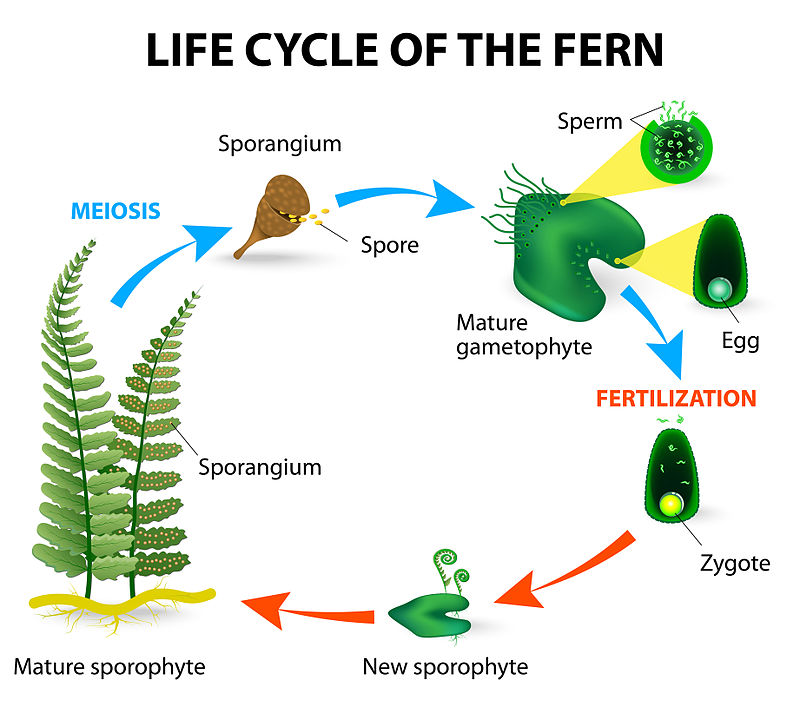
All non-vascular plants and most seedless vascular plants are homosporous, producing one type of spore (either male or female), giving rise to the gametophyte via mitosis. Certain genera of clubmoss (i.e. Selaginella) within Lycopodiopsida, are sporophytic plants generate two different kinds of spores (heterospory) producing separate male and female gametophytes. The gametophyte of heterosporous lycophytes is highly reduced while remaining enclosed in the sporangium plant. Once the sporophyte matures, it will develop multicellular structures, known as a megasporangium. Inside the megasporangium, is an immature egg cell, known as a megasporophyte. This cell undergoes meiosis producing four haploid egg cells, known as megaspores. Once meiosis occurs this structure is known as a megagametophyte. Megagametophytes are large multi-cellular structures producing and housing four megaspores (or eggs), while microgametophytes are much smaller, multi-cellular structures producing and housing sperm. Microgametophytes stem from diploid microsporangia, in which the microsporocytes within the microsporangium undergo meiosis producing haploid microspores.
Most seedless vascular plants (including homosporous members of Lycopodiopsida) have separate gametophytic and sporophytic forms. While these forms may be physically attached to each other, lycophytes that produce megagametophytes and microgametophytes, generate highly reduced gametophytes enclosed within the sporophytic tissue, seen in the micro- and megagametophytes. Once fertilization of the egg occurs, diploid tissue of the archegonium continues to grow enveloping the sporophytic embryo, developing into a mature megagametophyte. The mature megagametophyte is surround by a single epidermal layer of cells known as a megaspore wall. Once the fertilized megagametophyte germinates, the developing embryo is supplemented with food stored within the infertile tissue of the megagametophytes, similar to seed plants. Other seedless vascular plants do not have this food reserve, providing these lycophytes a competitive advantage over other seedless vascular plants during the early stages of development.
.png)
Gymnosperm life cycle
Gymnosperms have naked seeds
Extant seed plants (or spermatophytes) include gymnosperms and angiosperms. Fossil evidence suggests that seed plants originated from now extinct seed ferns. Gymnosperms (Greek for “naked seeds”) are the most primitive, extant seed plants and produce seeds that are unenclosed, while angiosperms (Greek for “vesseled seeds”) are flowering plants, which have their seeds encased (or vesseled) in an additional tissue, known as the fruit. Extant gymnosperms include cycads (Subclass Cycadidae), gingko tree (Subclass Ginkgoidae), gnetophytes (Subclass Gnetidae) and conifers (Subclass Pinidae).
Through the evolutionary trajectory of land plants, there has been a general trend towards a reduction of the gamete. Seed plants take this trend to an extreme. Bryophytes are gamete dominant. Seedless vascular plants are sporophyte dominant, yet their gametophytes are visible to the naked eye. Seed plant gametophytes are further reduced to microscopic proportions. Gametophytes in seed plants develop from sporangia and are retained within sporophytic tissues. Enveloping the gametophyte in sporophytic tissue is advantageous for land plants, as it protects the gametophyte from drying out. Seed plants are exclusively heterosporous, producing two different types of spores. Similar to heterosporous lycophytes, seed plants have highly reduced gametophytes. The microsporangium is a structure that houses microsporocytes. The diploid microsporocytes undergo meiosis producing haploid microspores. The microspores undergo mitosis, giving rise to the male gametophyte, or microgametophyte. The microgametophyte in seed plants is a four-celled haploid structure known as pollen.
Megagametophytes in lycophytes consist of a sporophytic (2n) embryo surrounded by nutritive tissue, which is all encased in a megaspore wall. The megasporangium is a diploid, multicellular structure that is composed of the megasporagium (known as the nuclleus in seed plants), which houses a diploid megasporoctye (known as the megaspore mother cell in seed plants). The nuclleus eventually becomes food reserves for the developing embryo. Pollination occurs when a pollen grain comes in contact with the nuclleus. This causes the megaspore mother cell to undergo meiosis producing four, haploid megaspores. Three of the megaspores perish, leaving one megaspore. The surviving megaspore undergoes mitosis to give rise to the female, haploid gametophyte, the megagametophyte. The megagametophyte is a multicellular structure that houses an egg enveloped in a vase-shaped archegonium. Fertilization occurs when the sperm and egg cells fuse.
Unlike megagametophytes in certain lycophytes, megagametophytes have an additional tissue, an integument. These modified megagametophytes of seed plants are ovules. The integument develops into a hard seed coat. This extra protection allows seeds to remain dormant until suitable environmental conditions emerge before they germinate. Lycophytes that develop megagametophytes do not have a seed coat and are not capable of extended dormancy prior to germination. Once the egg is fertilized, it develops into a diploid sporophytic embryo, housed inside the megagametophyte, surrounded by diploid food reserves from the nuclleus. After fertilization, the integument encloses the nuclleus and developing embryo, and hardens to become a seed coat. Once the seed germinates it develops into a sporophytic (2n) plant, or sporophyte. Once the plant reaches sexual maturation, it develops micro- and megasporangia, beginning the life cycle again. In pines, these are housed in cones.
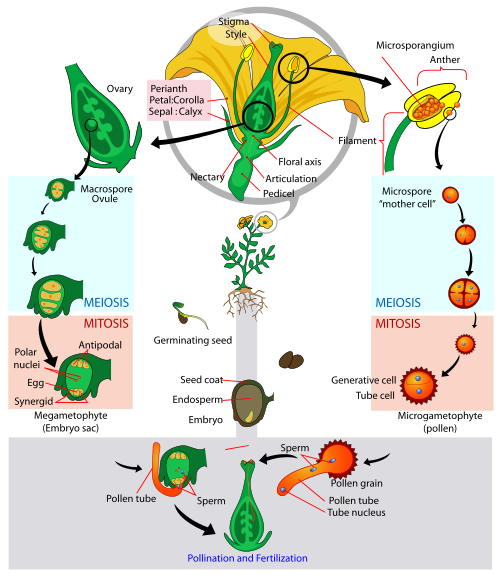
Angiosperm life cycle
Angiosperms have seeds enclosed in fruit
Angiosperms are the flowering plants. Angiosperms are presently the most dominant plants on the continents, represented by monocots (Class Monocotyledonae) and dicots (Class Dicotyledonae). Like all vascular plants, angiosperms are sporophyte dominant. Like gymnosperms, angiosperms are heterosporous, seed plants. Unlike gymnosperm, the seeds of angiosperms contain an additional tissue, known as the fruit. The fruit’s primary function is seed dispersal. Fruits can eaten by animals, stick to animals fur or feathers, or be carried by wind or water. Dispersing seeds away from the mother plant is beneficial for developing offspring as it ensures the offspring are not completing with the mother plant. Like gymnosperms, angiosperms produce microsporocytes that undergo meiosis to produce haploid microspores. Theses microspores subsequently divide via mitosis producing two-celled microgametophytes, known as pollen. Microsporocytes are held within a microsporangium, which is housed within a structure known as an anther. Flowers generally have many anthers, which are pollen-producing structures, which are typically yellow (sometimes orange) held on top of filaments (typically white) attached to the base of the flower.
Like gymnosperms, angiosperms produce ovules. However, gymnosperm ovules are said to be “naked.” Ovules in angiosperms are enclosed inside an additional tissue known as an ovary. Fruits develop from ovaries. The ovaries (typically housing many ovules) are a part of a larger structure known as a carpel. Atop the ovary extents a neck (or tube), known as a style. Atop the style, is an organ known as the stigma, which contains a sticky substance to hold pollen once it is deposited. The ovules inside the ovary contain a megasporangium housed within two layers of integuments. A diploid megasporocyte (or mother megaspore) within the megasporagium undergoes meiosis producing four haploid megaspores, of which only one survives. The surviving megaspore undergoes mitosis, producing an 8-celled female gametophyte, known as the embryo sac. Two cells closest to the equator of the embryo sac fuse producing a diploid polar nucleus. The remaining cells of the megagametophyte migrate away from the equator of the embryo sac. The three cells furthest from the opening of the ovule are known as antipodals, while the three remaining cells (closest to the opening of the ovule) include one egg cell and two synergid cells (or “helper” cells).
Once the pollen reaches the stigma, a pollen tube grows down the style and enters the ovary between the integuments. Two sperm cells are deposited in the embryo sac. Next, a unique process to angiosperms occurs, known as double fertilization. Like all other plants, one sperm and egg fuse creating a diploid zygote, which develops into an embryo. The second sperm fuses with a the diploid polar nucleus, forming a triploid (3n) cell that develops into a food reserve tissue for the developing embryo, known as the endosperm. Following fertilization, the integuments hardened forming a tough outer seed coat, similar to the gymnosperms. Once the seed germinates, a root radicle emerges followed by either one (in the case of monocots) or two (in the case of dicots) seed leaves, known as cotyledons. As the seedling develops it is supplemented with food reserves from the endosperm. Angiosperms can have stamens and pistils on the same flower (hermaphroditic), stamens and pistils on different flowers (monoecious) or separate male and female plants (dioecious). Nearly all flowering plants have built in mechanisms to reduce self-pollination, either by physical or temporal separation of stamen and pistol development.