
Chapter: A Brief History of Life
What Is a Phylogeny?
The word phylogeny comes from two roots: “phylo,” meaning branch, and “geny,” meaning origin or creation. Together, the term refers to the branching evolutionary history of life on Earth. More than just a definition, phylogeny represents a way of understanding how organisms are connected through evolution over time. Phylogenetics is the scientific study of these relationships. Using genetic data, physical characteristics, and fossil evidence, scientists reconstruct the evolutionary history of organisms and their relationships to one another. These relationships are then represented in diagrams called phylogenetic trees, which function much like evolutionary family trees. The closer two species, or taxa, are positioned on a phylogenetic tree, the more recently they shared a common ancestor and the more closely related they are. Scientists build phylogenetic trees by combining evidence from DNA comparisons, morphological analysis, and the fossil record. DNA provides molecular evidence of relatedness, while fossils offer direct evidence of ancient organisms, including what they looked like, when they lived, and where they existed. Together, these sources of evidence allow scientists to reconstruct the history of life with increasing accuracy.
Reading a Phylogenetic Tree
Understanding the structure of a phylogenetic tree is essential for interpreting evolutionary relationships among organisms. Branches represent the evolutionary path of a lineage through time. Each branch traces the history of a group of organisms as they evolve from common ancestors. Nodes are the points where branches split, representing speciation events in which one ancestral species diverged into two separate lineages. These branching points indicate a shared common ancestor. The tips of the phylogenetic tree (known as terminal nodes) are located at the ends of branches and represent either living or extinct species. Branches that extend to the present indicate species that are still alive today, while branches that end earlier on the tree represent lineages that became extinct.
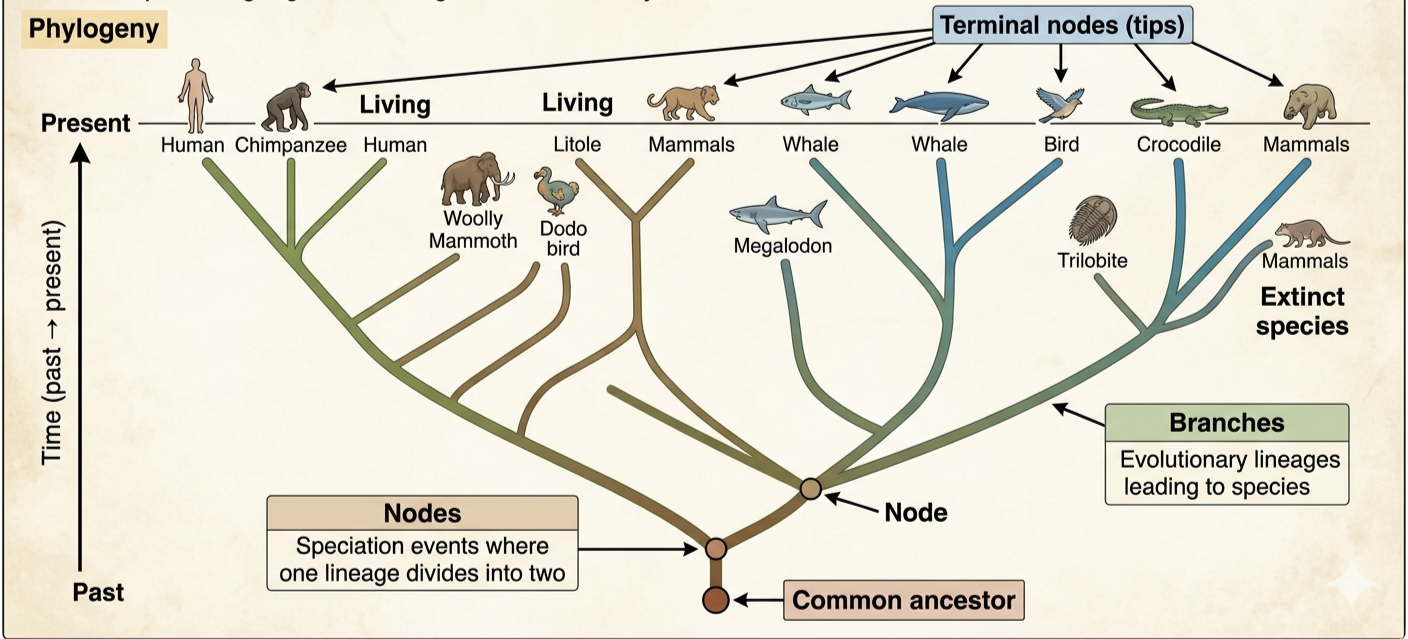
Figure 1. Reading Phylogenetic Trees. Phylogenies describe the branching evolutionary history of life. Phylogenetic trees represent these relationships using branches, nodes, and common ancestors. Branches trace lineages through time, nodes represent speciation events, and terminal nodes represent living or extinct species. Phylogenetic trees are constructed using evidence from DNA, fossils, and morphology to reveal how closely related organisms are based on shared evolutionary history.
Parsimony: Keeping Evolutionary Explanations Simple
Parsimony is a guiding principle used in constructing most phylogenetic trees. It holds that the simplest explanation, the one that requires the fewest evolutionary changes, is usually the most likely to be correct. In phylogenetics, this means favoring evolutionary scenarios that involve the smallest number of changes rather than assuming that complex features evolved multiple times independently. By doing this, scientists can infer the most likely evolutionary relationships among organisms. For example, if you compare several shapes where three are circles and one is a triangle, parsimony would group the three circles together as more closely related because they share the same traits, while the triangle would represent a separate lineage.
However, evolution does not always follow the simplest path. Similar traits can evolve independently in unrelated groups, which means that the true evolutionary history can sometimes be more complex than what the most parsimonious explanation suggests (see homoplasy below). Before we get into the complexity, let’s start at the beginning.
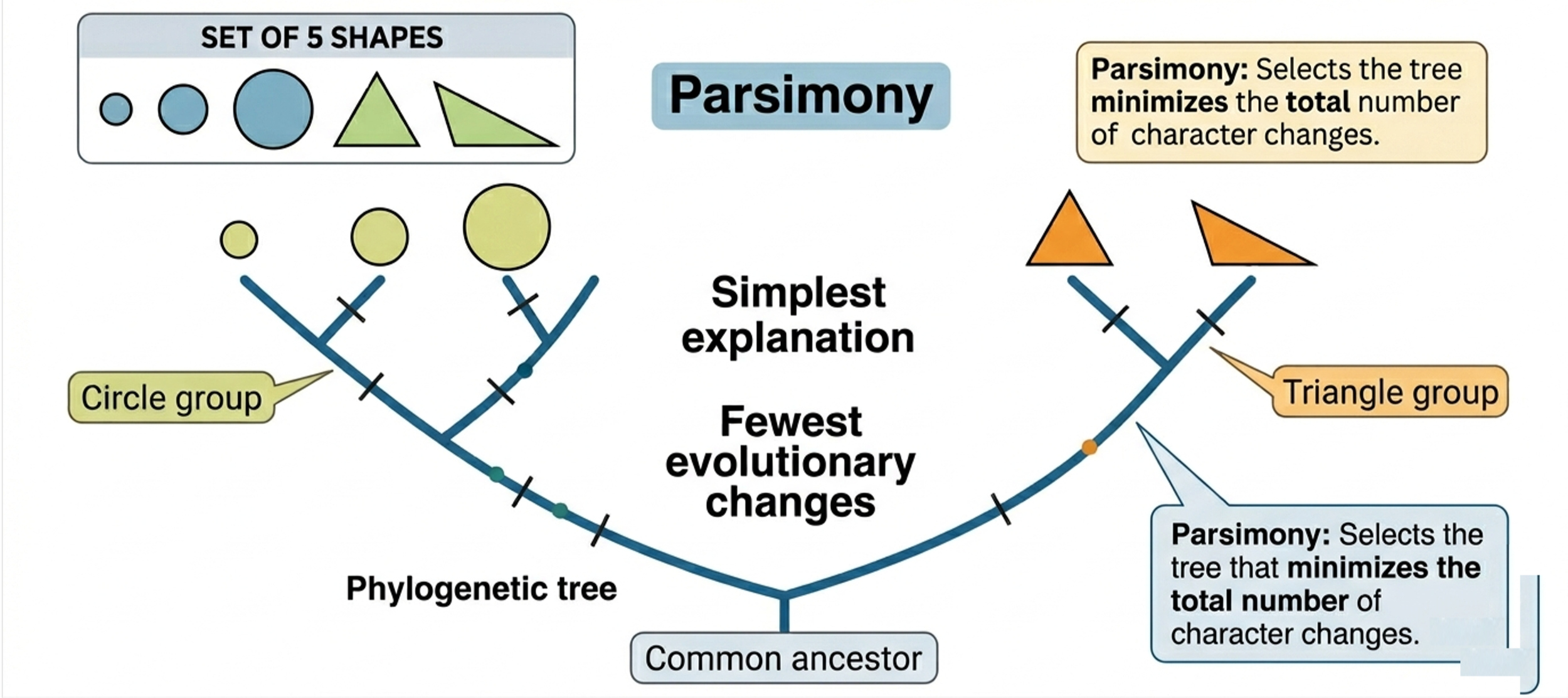
Figure 2. Parsimony. Parsimony is a guiding principle in phylogenetics that favors the simplest explanation requiring the fewest evolutionary changes. It is used to construct phylogenetic trees by grouping organisms in ways that minimize the number of changes needed to explain their relationships, helping scientists infer the most likely evolutionary history.
How Phylogenies Are Built
Scientists build phylogenetic trees using two main approaches: phenetics and cladistics. Using parsimony as a guiding principle, both aim to infer evolutionary relationships based on similarity. Both assume physical similarity infers evolutionary similarity. They differ in their measures of physical similarity. The phenetic approach compares similarity at a molecular scale, using genetic similarity (i.e. DNA) to determine evolutionary relationships. The cladistic approach determines evolutionary relationships based on morphological characteristics (i.e. fossil structure).
Both approaches create clades, in which organisms with more similarities are placed closer together on the phylogenetic tree and are considered more closely related than organisms with more different genetic sequences. Clades exist at different hierarchical scales. A species is the smallest basic unit, representing a single lineage capable of interbreeding. Closely related species are grouped into a genus, genera are grouped into families, and this pattern continues upward into orders, classes, phyla, and kingdoms. Each higher level represents a broader clade that includes more distant shared ancestry, while lower levels reflect more recent divergence. For example, multicellular animals form a large clade. Within that group, animals with body symmetry form a more specific clade. That group can be further divided into lineages with bilateral symmetry and those with radial symmetry. Each step reflects a narrower set of shared derived traits and a more recent common ancestor.
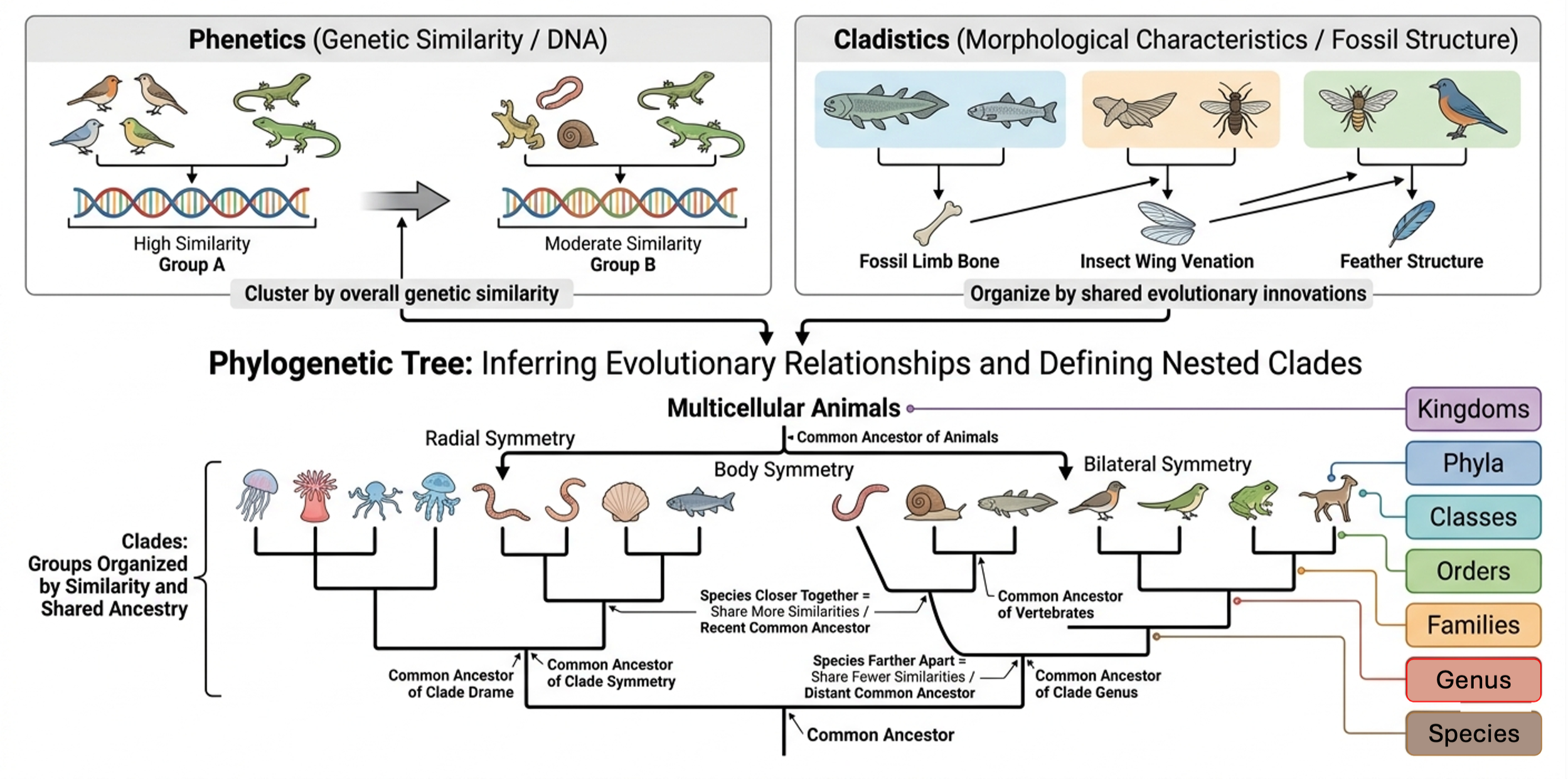
Figure 3. How Phylogenies are Built. Scientists build phylogenetic trees using phenetics, which compares genetic similarity, and cladistics, which uses morphological characteristics. Both approaches infer evolutionary relationships based on similarity and produce clades in which organisms with more similarities are grouped more closely together. These clades exist at multiple hierarchical levels, from species up through broader groups such as genus, family, and higher taxonomic levels, with each level representing different degrees of shared ancestry.
The Phenetic Approach
The phenetic approach groups organisms based on overall similarity, most often using comparisons of DNA sequences, and less commonly RNA or protein data.Organisms with more similar genetic sequences are placed closer together on the tree and are considered more closely related than organisms with more different genetic sequences. This method is fast, quantitative, and well-suited for large datasets. However, it focuses on similarity alone and does not distinguish why organisms look alike, meaning it can sometimes group organisms together for reasons other than shared ancestry.
The Cladistic Approach
The cladistic approach reconstructs evolutionary relationships by comparing physical morphological characteristics, meaning observable traits such as body structure, symmetry, and anatomical features. It focuses specifically on shared derived traits called synapomorphies, which are characteristics that evolved in a common ancestor and were passed down to all its descendants. These synapomorphies form the basis for building phylogenetic trees because they reveal patterns of shared ancestry. Organisms are grouped together when they share these derived traits, and each synapomorphy helps define branching points where lineages split. In this way, cladistics builds a tree of life by using inherited physical characteristics to map evolutionary relationships and progressively organize organisms into increasingly specific clades. In this way, cladistics works like reconstructing a branching history: scientists move from broad shared traits at higher taxonomic levels down to more specific, recently diverged groups, using inherited characteristics to map how life has diversified over time.
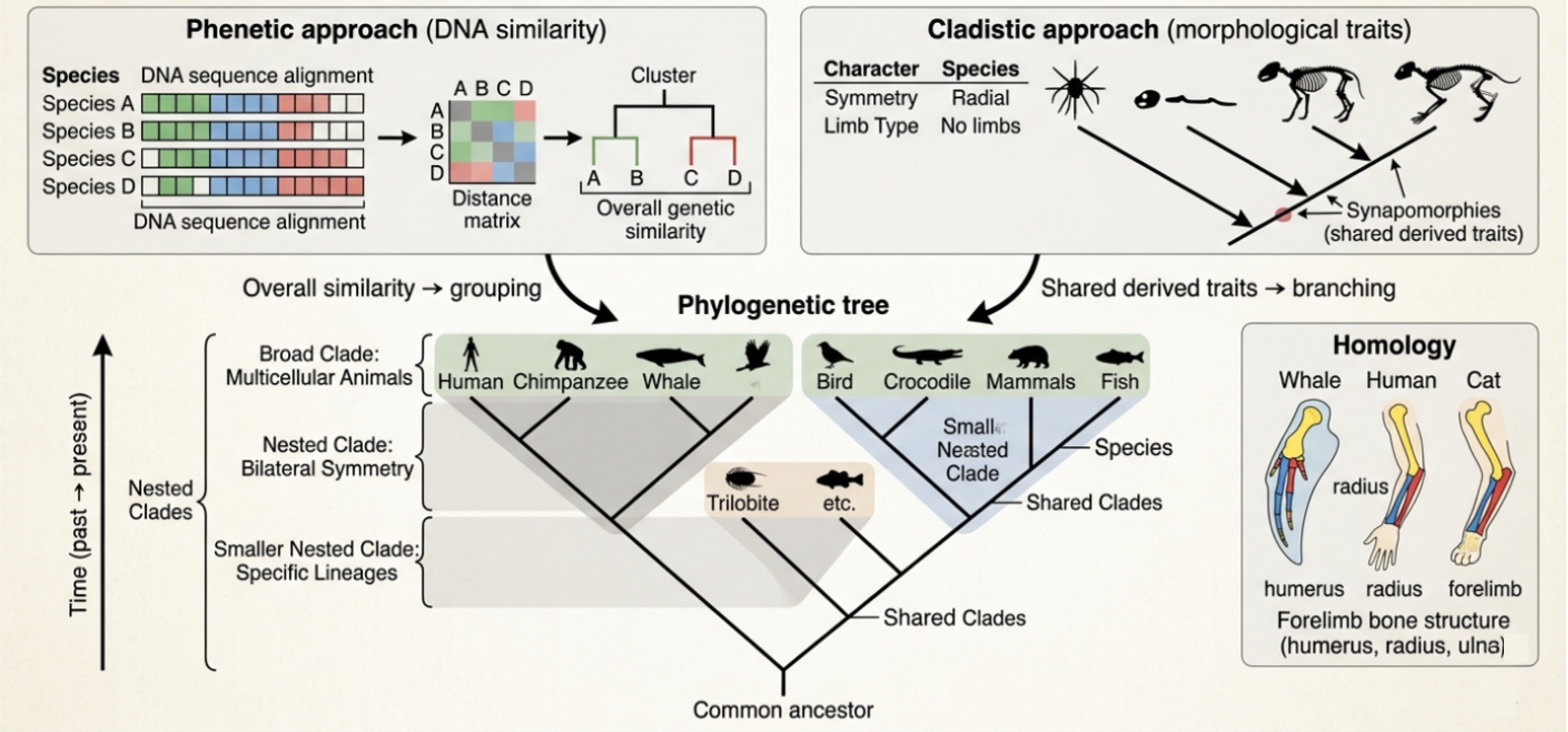
Figure 4. Building Phylogenetic Trees. Phylogenetic trees are constructed using both phenetics, which groups organisms based on genetic similarity, and cladistics, which uses shared derived morphological traits to infer evolutionary relationships. These approaches help identify clades, or groups of organisms that share common ancestry at different hierarchical levels. Evidence from both DNA and anatomy, including homologous structures such as vertebrate forelimbs, improves the accuracy of reconstructing evolutionary history.
Genetics and Anatomy Together Reveal Evolutionary Relationships
Phenetics and cladistics work together to improve the accuracy of phylogenetic trees by separating true evolutionary similarity from misleading resemblance. Phenetics organizes organisms based on overall genetic similarity, while cladistics evaluates relationships using shared derived traits to infer common ancestry. A key concept here is homology, where similarities arise from shared ancestry. For example, whales, cats, and humans all share the same underlying forelimb bone structure, even though those limbs are adapted for very different functions such as swimming, walking, and grasping. Despite these differences in appearance and use, the bones follow a consistent anatomical pattern (humerus, radius, ulna, and associated hand bones) across all three groups. This shared structural design is evidence of homology, meaning it was inherited from a common ancestor rather than independently evolved. It shows that evolution modifies existing structures over time instead of starting from completely new designs in each lineage.
Homoplasy and Convergent Evolution
In contrast, homoplasy refers to traits that look similar but evolved independently due to similar environmental pressures, a process known as convergent evolution. For example, dolphins and ichthyosaurs both evolved streamlined bodies, fins, and sharp teeth for efficient swimming, which can make them appear closely related in a phenetic sense. However, cladistic and deeper anatomical analysis separates them, placing dolphins within mammals based on features such as mammary glands, lung-based respiration, and a mammalian skeletal structure inherited from a terrestrial ancestor, while ichthyosaurs remain within reptilian lineages. Phylogeneticists use this combined evidence to distinguish homology from homoplasy and build trees that reflect true evolutionary history rather than surface-level similarity. To tell homology from homoplasy, scientists look for intermediate forms and genetic evidence. Homologous traits usually show up in transitional fossils and share common genetic signatures.
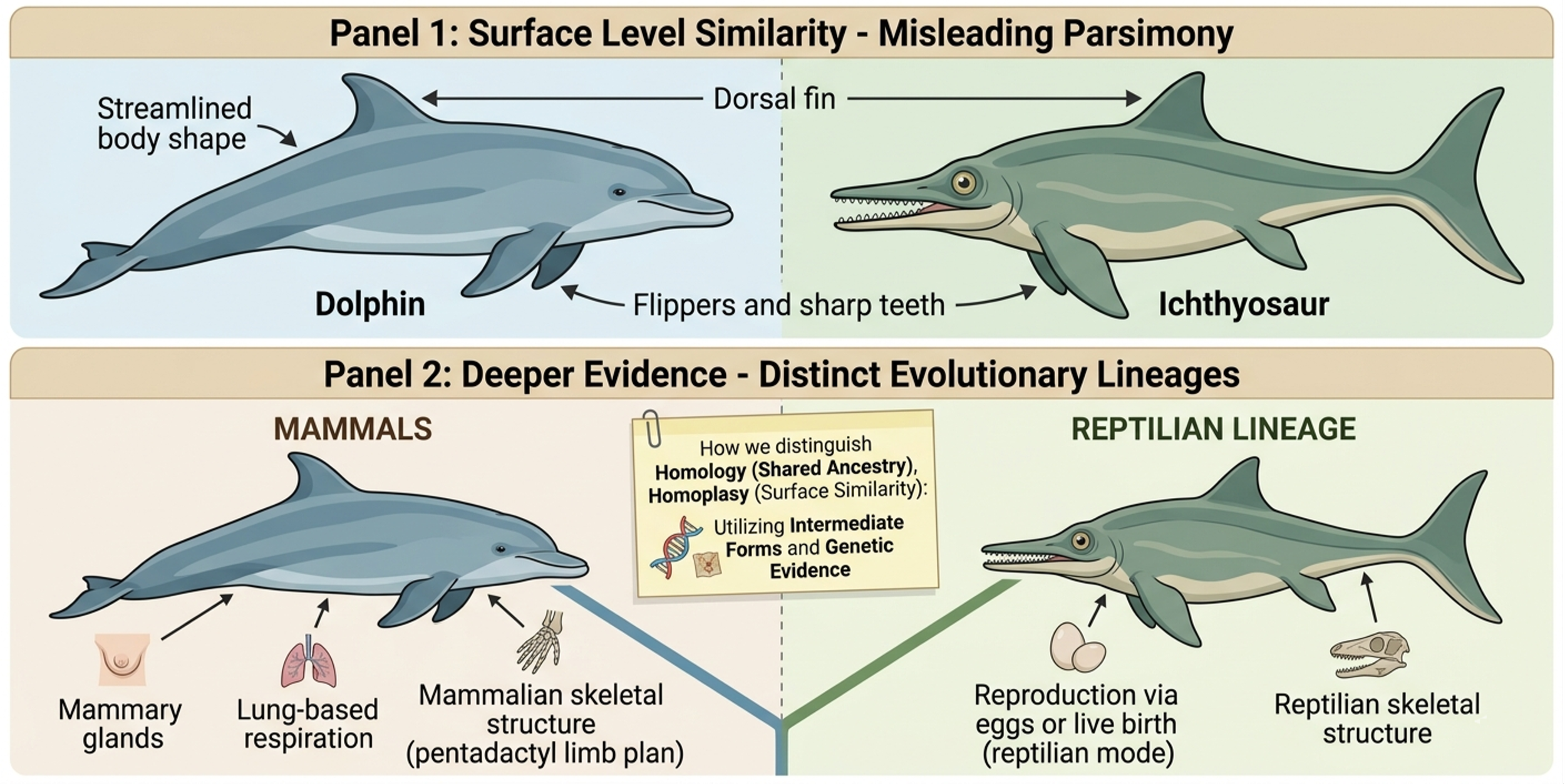
Figure 5. Homoplasy and Convergent Evolution. Homoplasy refers to similarities that evolve independently due to similar environmental pressures, a process known as convergent evolution. Dolphins and ichthyosaurs share similar body forms adapted for swimming, but deeper anatomical and genetic evidence places them in separate evolutionary lineages. Scientists distinguish homology from homoplasy using anatomical traits, fossils, and genetic data to reconstruct true evolutionary history.
When Parsimony Fails: Whale Evolution
Whales are genetically most closely related to hippos, which belong to the artiodactyls (even-toed hoofed mammals such as cows, pigs, and deer). A defining feature of artiodactyls is a specialized ankle bone called the astragalus, which helps identify the group morphologically. However, whales lack this bone because they lost it during their transition back to an aquatic lifestyle, meaning their anatomy no longer fits the typical artiodactyl pattern. If classification relied only on morphology, whales would appear to fall outside this group, despite strong structural similarities among other artiodactyls. Genetic evidence shows a different picture: whales and hippos share a more recent common ancestor with each other than with any other living mammals. To reconcile this, evolutionary history must account for trait loss—recognizing that the astragalus was present in the common ancestor of artiodactyls and secondarily lost in whales. This produces a less intuitive but more accurate phylogenetic explanation supported by both DNA and evolutionary reasoning.
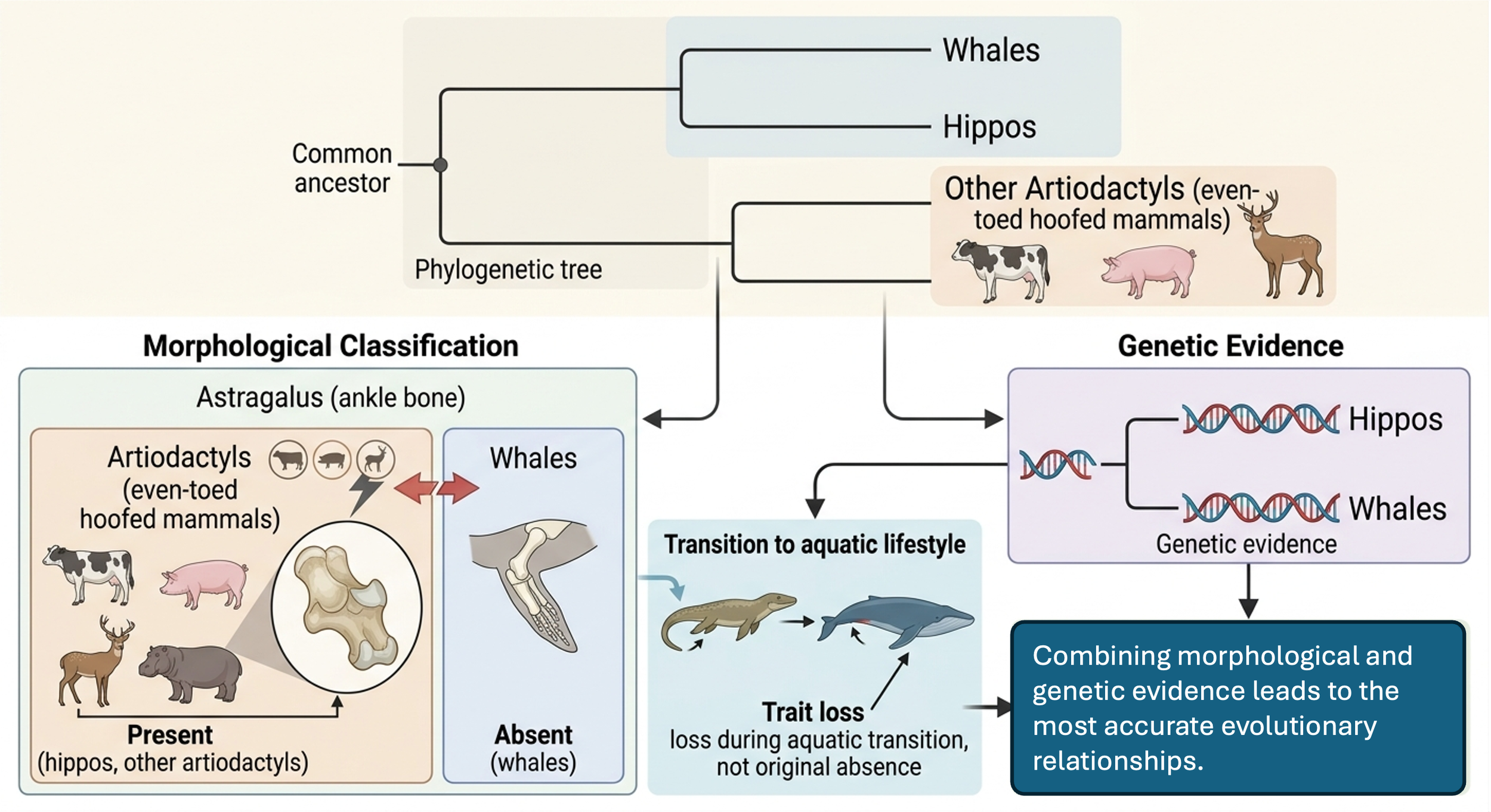
Figure 6. Whale Evolution. Whales are genetically most closely related to hippos, even though morphological classification based on traits such as the astragalus bone would not place them together. Whales lost this bone during their transition to an aquatic lifestyle, demonstrating that trait loss can obscure evolutionary relationships. Genetic evidence reveals their closer common ancestry, showing that accurate phylogenies must account for both DNA and evolutionary changes in traits over time.
A Brief History of Life on Earth
Dividing Earth’s History
To organize Earth’s vast and complex history, geologists divide geologic time into a hierarchical system of units that allows them to describe events across billions of years in a structured way. The largest divisions are called eons, which represent the broadest spans of time in Earth’s history and encompass major changes in the planet’s atmosphere, geology, and life. Each eon is further subdivided into eras, which mark significant shifts in dominant life forms and global conditions. Eras are then divided into even smaller units called periods, which capture more specific intervals defined by distinct biological and geological events recorded in the rock and fossil record. This hierarchical system provides a consistent framework for understanding how Earth has changed over time from its formation to the present.
Earth’s history is divided into four major eons, which represent the largest units of geologic time and capture the most fundamental transitions in the planet’s development. The Hadean Eon (about 4.6 to 4.0 billion years ago) begins with Earth’s formation. During this time, the planet was extremely hot, largely molten, and constantly bombarded by space debris. No known life existed, and the surface was unstable as the crust, atmosphere, and early oceans began to form. The Archean Eon (about 4.0 to 2.5 billion years ago) marks the stabilization of Earth’s crust and the appearance of the earliest known life. These first organisms were simple, single-celled prokaryotes that lived without oxygen in the atmosphere, which was still very different from today’s oxygen-rich environment. The Proterozoic Eon (about 2.5 billion to 541 million years ago) saw major biological and atmospheric changes, including the rise of oxygen through photosynthesis and the development of more complex eukaryotic cells. By the end of this eon, multicellular life began to emerge, setting the stage for more complex ecosystems. The Phanerozoic Eon (541 million years ago to present) is the current eon and is defined by the explosion of visible, complex life in the fossil record. It includes the diversification of animals, plants, and fungi, as well as all major groups of life seen today, including the rise of humans.
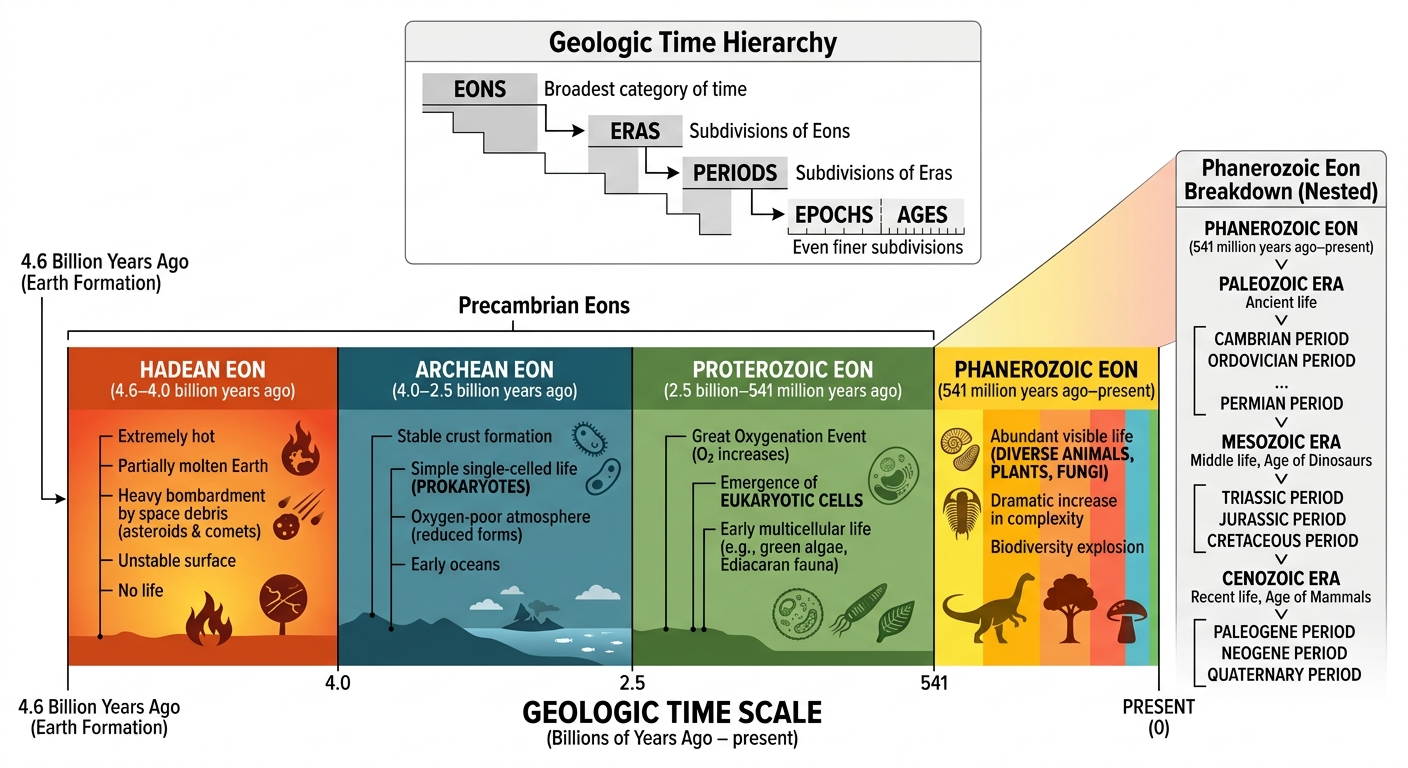
Figure 8. Earth’s Geologic Timeline. Geologic time is organized into a hierarchical system of eons, eras, and periods to describe Earth’s history over billions of years. The four major eons—Hadean, Archean, Proterozoic, and Phanerozoic—represent major transitions from Earth’s formation, to early life, to increasing oxygen and complexity, and finally to the diversification of complex life forms seen today.
The Hadean Eon (4.6 – 4.0 Billion Years Ago)
The Hadean Eon (4.6 – 4.0 billion years ago) represents the earliest chapter in Earth’s history, beginning with the formation of the solar system and the assembly of the planets from the solar nebula. This nebula was a rotating cloud of gas and dust enriched by heavy elements produced in earlier generations of stars that ended in supernova explosions. As gravity caused part of this cloud to collapse, it formed a spinning protoplanetary disk in which the Sun formed at the center, while surrounding material gradually accreted into planetesimals and ultimately the planets, including Earth. Hadean Earth was defined by extreme conditions, including widespread volcanism, intense bombardment by asteroids and planetesimals, and constant reshaping by internal heat and external impacts. As a result, the planet had no stable surface in the modern sense, and its environment was dominated by rapid physical and chemical transformation.
Planetary Formation and Early Catastrophic Events
In its earliest stages, Earth grew through continuous collisions with surrounding space debris, which released enormous amounts of energy and maintained a largely molten surface. One of the most significant events of this eon was a giant impact between Earth and a Mars-sized body, which ejected material into orbit that eventually coalesced to form the Moon. This impact reshaped Earth’s internal structure, influenced its rotational dynamics, and helped stabilize its axial tilt over long timescales, an important factor in the development of long-term climate stability.
Cooling, Crust Formation, and Early Atmosphere
As the Hadean progressed, Earth gradually cooled enough for the first thin and unstable crust to form. However, this early crust was repeatedly destroyed and recycled by ongoing impacts, widespread volcanism, and internal melting driven by radioactive decay. Volcanic outgassing released large volumes of gases into the atmosphere, forming a primitive atmosphere composed mainly of water vapor, carbon dioxide, nitrogen, and other volcanic compounds. While this atmosphere played a critical role in shaping early planetary chemistry, it would have been extremely harsh and inhospitable by modern biological standards.
Formation of the First Oceans and Early Earth Conditions
As temperatures continued to decline, water vapor in the atmosphere condensed and accumulated on the surface, leading to the formation of Earth’s earliest oceans. This transition marked a major turning point in Earth’s evolution, as the planet shifted from a fully molten state to one with persistent liquid water and stable surface environments. These early oceans provided the first settings capable of supporting complex chemical interactions, laying the groundwork for prebiotic chemistry. Although no confirmed fossil evidence of life exists from the Hadean Eon, this interval established the essential physical and chemical conditions required for life to emerge in the following Archean Eon, including liquid water, energy gradients, and chemically active environments.
Dating the Early Earth and Geological Evidence
Because Earth is geologically active, much of its earliest crust has been recycled through plate tectonics, melting, and erosion, making direct evidence from the Hadean extremely rare. Scientists reconstruct this period using radiometric dating techniques, which measure the decay of radioactive isotopes in minerals and meteorites. Methods such as potassium-argon dating consistently indicate an age of approximately 4.6 billion years for the solar system. The oldest known terrestrial rocks, found in regions such as northern Quebec, date to about 4.0 billion years ago and provide rare, indirect windows into Earth’s earliest crustal history. Together, these lines of evidence form the foundation for understanding the origin and early evolution of the Earth system.
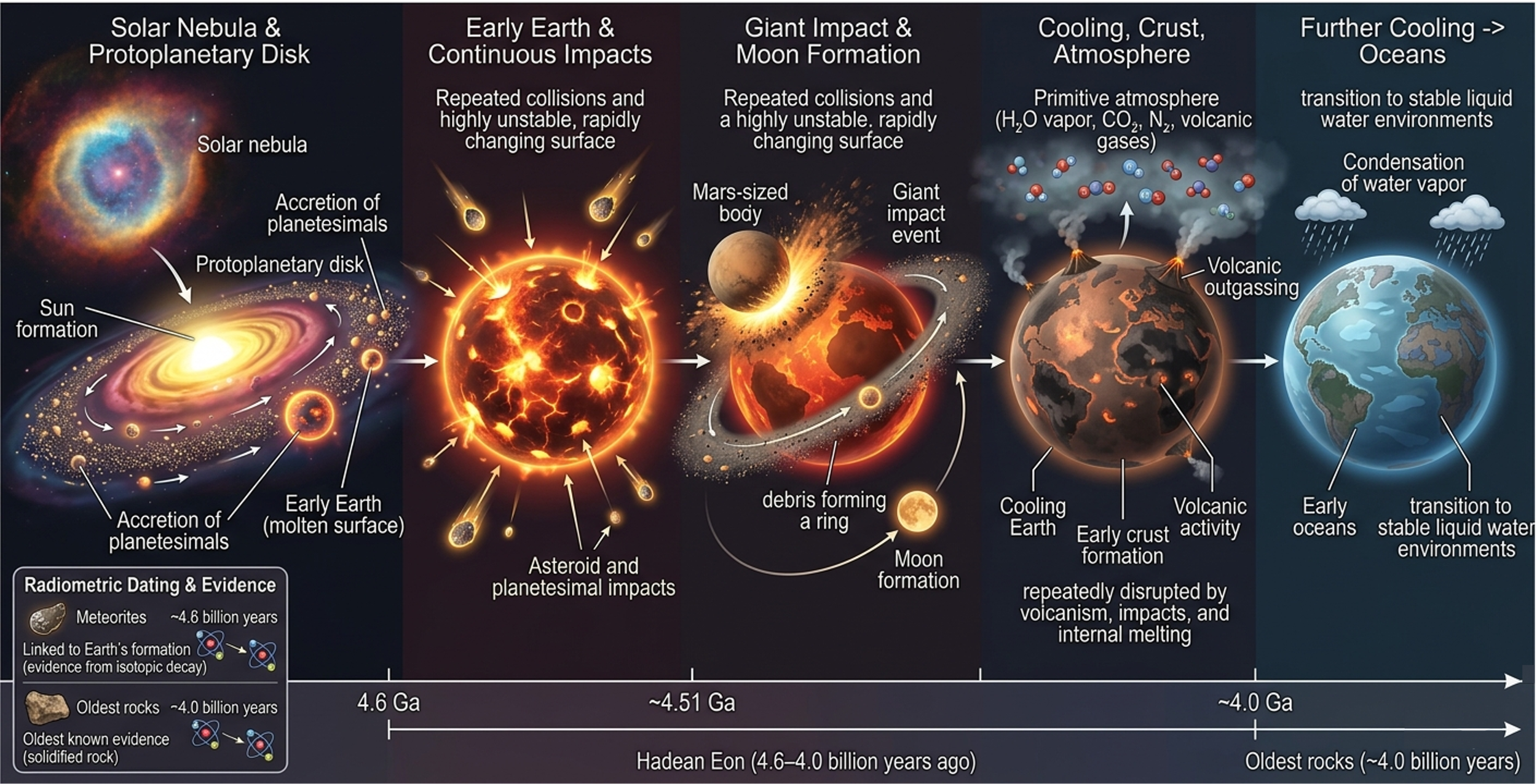
Figure 7. Birth of Earth. The Hadean Eon represents the earliest stage of Earth’s history, beginning with the formation of the solar system from a collapsing solar nebula. Early Earth formed through accretion, remained largely molten due to intense impacts, and experienced a giant impact that formed the Moon. As the planet cooled, a thin crust and primitive atmosphere formed, followed by the condensation of water vapor and the creation of early oceans. Radiometric dating of meteorites and ancient rocks provides evidence that Earth formed about 4.6 billion years ago.
The Archean Eon (4.0 – 2.5 Billion Years Ago)
The Archean Eon (4.0 – 2.5 billion years ago) marks a major transition in Earth’s history as the planet cooled enough for a stable crust and early oceans to form, creating more favorable conditions for life. During this time, Earth’s surface shifted from a highly volatile, impact-driven environment to one where solid continental crust began to persist for longer periods. These early oceans provided a stable liquid water environment that allowed chemical processes essential for life to develop and persist. Although oxygen was largely absent from the atmosphere, energy sources such as volcanic activity, hydrothermal vents, and ultraviolet radiation likely drove the chemical reactions that supported the earliest forms of metabolism.
First Evidence of Life: Chemical
Evidence for early life is found in rocks from Greenland dated to about 3.85 billion years ago, which show an unusual enrichment of carbon-12, an isotope that living organisms preferentially use during biological processes such as photosynthesis and metabolism. This enrichment is identified by measuring the ratio of carbon isotopes in ancient mineral samples and comparing it to what would be expected from purely non-biological processes. Living organisms preferentially use the lighter carbon isotope (carbon-12) over carbon-13 during processes like photosynthesis and metabolism, creating a measurable carbon-12 enrichment, while non-biological processes do not strongly favor one isotope and therefore produce more balanced isotope ratios. The pattern suggests biological activity was influencing carbon cycling at the time, making it one of the earliest chemical signatures potentially linked to life on Earth.
First Evidence of Life: Physical
The first widely recognized fossil evidence of life appears in stromatolites, which are layered rock structures formed by microbial communities, primarily cyanobacteria, that trapped and bound sediments in shallow marine environments. These microbial mats grew in thin layers over time, repeatedly capturing fine particles and minerals, which gradually built up distinctive dome-shaped structures that preserve a physical record of early biological activity. Because stromatolites form through the interaction of living organisms with their environment, they provide both structural and behavioral evidence of some of the earliest ecosystems on Earth. Fossilized stromatolites dating to about 3.45 billion years ago have been discovered in Western Australia, offering strong evidence that life was already established and widespread in shallow seas, while modern stromatolites still exist today in limited, protected environments such as Shark Bay, Australia.
Life in the Archean Eon
Archean Life was simple, microscopic, and dominated Earth’s oceans long before complex organisms existed. The earliest life forms were unicellular prokaryotes, meaning they lacked a nucleus and other internal structures, and they likely resembled modern bacteria and archaea in basic form. These organisms lived in an anoxic environment, where oxygen was essentially absent, so they relied on anaerobic metabolic pathways or early forms of chemosynthesis to obtain energy. Many early Archean organisms likely lived near hydrothermal vents or in shallow marine environments, where chemical-rich fluids provided a steady energy source and protection from harsh surface conditions like radiation and frequent impacts. These stable aquatic settings supported some of the earliest metabolic systems on Earth.
The Great Oxygenation Event
Toward the later Archean Eon, cyanobacteria evolved and began performing photosynthesis, using sunlight to produce energy and releasing oxygen as a byproduct. Although most oxygen was initially absorbed by rocks and oceans, this process slowly began transforming Earth’s atmosphere and set the stage for major changes in the Proterozoic Eon. The transition from the Archean to the Proterozoic Eon (around 2.5 billion years ago) is defined by one of the most important shifts in Earth’s history: the gradual buildup of oxygen in the atmosphere, known as the Great Oxygenation Event. During the late Archean, early photosynthetic organisms—especially cyanobacteria—began producing oxygen as a byproduct of photosynthesis, but most of it was initially absorbed by dissolved iron and other reactive materials in the oceans and crust. Once these oxygen “sinks” became saturated, free oxygen began accumulating in the atmosphere, dramatically altering Earth’s chemistry and climate. This transition also marks a major ecological turning point, as oxygen was toxic to many existing anaerobic organisms, forcing evolutionary adaptation, extinction, or retreat into oxygen-free environments. At the same time, the rising oxygen levels enabled the evolution of more energy-efficient aerobic metabolism, setting the stage for the development of more complex eukaryotic cells during the Proterozoic.

Figure 8. The Archaean Eon. The Archean Eon marks the stabilization of Earth’s crust and the formation of early oceans, creating conditions suitable for the first life. Evidence for early life includes carbon isotope signatures and fossil stromatolites formed by microbial communities in shallow seas. Life was dominated by simple prokaryotes living in oxygen-poor environments, until cyanobacteria began producing oxygen through photosynthesis, gradually transforming Earth’s atmosphere and setting the stage for major biological changes in later eons.
The Proterozoic Eon (2.5 Billion – 541 Million Years Ago)
The Proterozoic Eon (2.5 billion – 541 million years ago) was a long interval of major transformation in Earth’s biological and geological systems. Following the Great Oxygenation Event, oxygen continued to accumulate in the atmosphere and oceans, fundamentally altering Earth’s chemistry and enabling new forms of metabolism and increasingly complex life.
Continuing Oxygenation
One of the most important changes of the Proterozoic was the continued rise of oxygen produced by cyanobacteria. This oxygen reacted with dissolved iron in the oceans, forming widespread banded iron formations, which serve as geological evidence of shifting atmospheric and ocean chemistry over time. As oxygen became more available and stabilized in the environment, it enabled the evolution of aerobic respiration, a far more efficient metabolic process than anaerobic pathways, allowing organisms to extract more energy from food (in the form of ATP). Such a dramatic rise in efficiency rapidly accelerating natural selection, and therefore enhancing biological complexity.
Emergence of Eukaryotic Cells
The emergence of eukaryotes represents one of the most consequential transitions in the history of life, marked by the origin of both the nucleus and mitochondria and the dramatic expansion of biological complexity that followed. The nucleus likely evolved from infoldings of the plasma membrane that increasingly specialized into a protected compartment for genetic material, separating transcription from translation. This separation provided major advantages: it allowed tighter regulation of gene expression, enabled more complex RNA processing such as splicing, and reduced interference between transcription and protein synthesis, all of which supported larger genomes and more sophisticated cellular control systems. The origin of mitochondria, explained by endosymbiosis with an ancestral aerobic bacterium, provided an even more decisive energetic breakthrough. By converting oxygen into highly efficient ATP production, mitochondria vastly increased the energy available per gene and per cell, breaking the energetic constraints that had limited prokaryotic complexity. Together, these innovations created cells capable of greater size, internal compartmentalization, and regulatory complexity. The result was not just a new cell type but a new evolutionary platform, opening ecological opportunities that triggered extensive evolutionary radiation.
Increasing Biological Complexity
The Proterozoic Eon witnessed a profound shift in the trajectory of life on Earth, not only through the origin of multicellular organisms such as simple algae, but also through foundational biological innovations that reshaped how evolution operated. Multicellularity marked a critical departure from the long dominance of single-celled life, introducing coordinated cellular systems in which individual cells could specialize and function as parts of a larger, integrated organism. This allowed for new levels of biological organization, including the development of simple tissues, increased size, and the ability to exploit new ecological niches that were inaccessible to single cells. At the same time, evidence suggests that sexual reproduction emerged during this interval, dramatically altering evolutionary dynamics by introducing genetic recombination as a routine process. Unlike asexual reproduction, which produces genetically identical offspring, sexual reproduction generates novel gene combinations each generation, increasing genetic variation within populations and accelerating the pace of natural selection. Together, multicellularity and sex created a powerful evolutionary feedback loop: greater variation supplied the raw material for selection, while higher organizational complexity allowed organisms to respond to environmental pressures in more flexible and efficient ways. These innovations collectively set the stage for the later explosion of eukaryotic diversity, enabling life to become larger, more complex, and ecologically more dynamic than anything that had existed before.
Environmental Change and Glaciation
Toward the late Proterozoic, Earth experienced severe “Snowball Earth” glaciations in which ice may have covered much of the planet, potentially reaching near the equator. These conditions drastically reduced habitable environments, likely forcing life into limited refuges such as thin-ice zones, deep ocean vents, and isolated pockets of liquid water. Although extreme, these stresses created strong selective pressures and population bottlenecks that may have driven evolutionary innovation, favoring organisms with greater environmental tolerance and metabolic flexibility. As the planet warmed, ice retreated and more stable, oxygen-rich oceans reemerged. Higher oxygen levels supported more active and larger organisms, while expanding shallow marine environments encouraged increasingly complex ecosystems. By the end of the Proterozoic, these changes had established the environmental and biological foundations for the rapid diversification of life that followed in the Phanerozoic Eon.
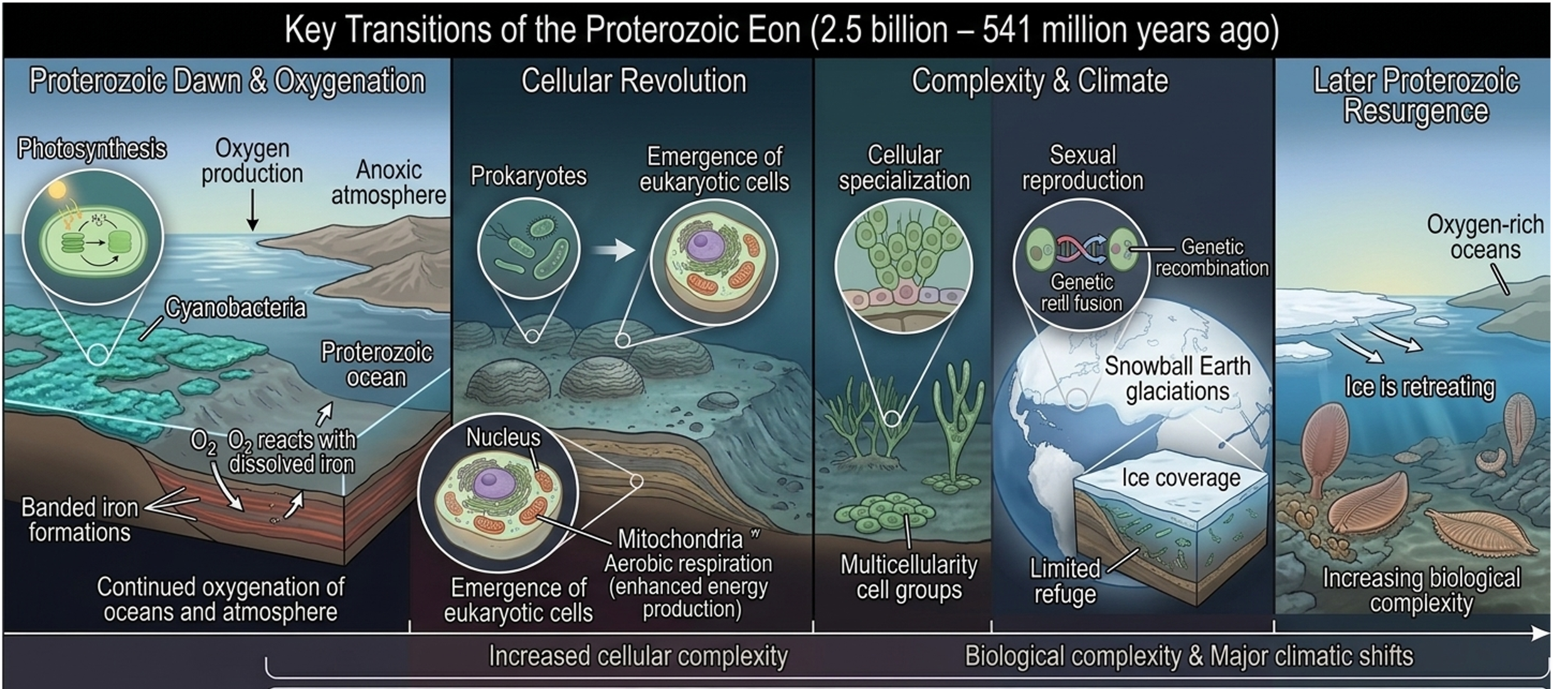
Figure 9. Proterozoic Eon. The Proterozoic Eon was a period of major transformation in Earth’s systems, marked by rising oxygen levels, the evolution of eukaryotic cells with internal complexity, and the emergence of multicellular life and sexual reproduction. These changes increased biological energy availability and genetic variation, driving greater evolutionary complexity. Severe global glaciations temporarily reduced habitable environments, but subsequent warming and ocean expansion set the stage for the diversification of complex life in later eons.
The Phanerozoic Eon (541 Million Years Ago – Present)
The Phanerozoic Eon marks the interval of Earth’s history during which complex multicellular life became widespread and abundant in the fossil record. Building on the environmental and biological changes of the Proterozoic, this eon saw the rapid diversification of plants, animals, and fungi, as well as the development of increasingly complex ecosystems on land and in the oceans. The Phanerozoic is divided into three major eras: the Paleozoic, Mesozoic, and Cenozoic. The Paleozoic Era included the Cambrian Explosion and the movement of life onto land. The Mesozoic Era was dominated by reptiles and dinosaurs, while the Cenozoic Era saw the diversification of mammals, birds, and modern ecosystems following the extinction of the non-avian dinosaurs. Throughout the eon, plate tectonics, climate change, evolutionary competition, and mass extinctions continually reshaped Earth’s environments and ecosystems. These changes repeatedly created new evolutionary opportunities, driving the ongoing diversification and adaptation of life across geological time.
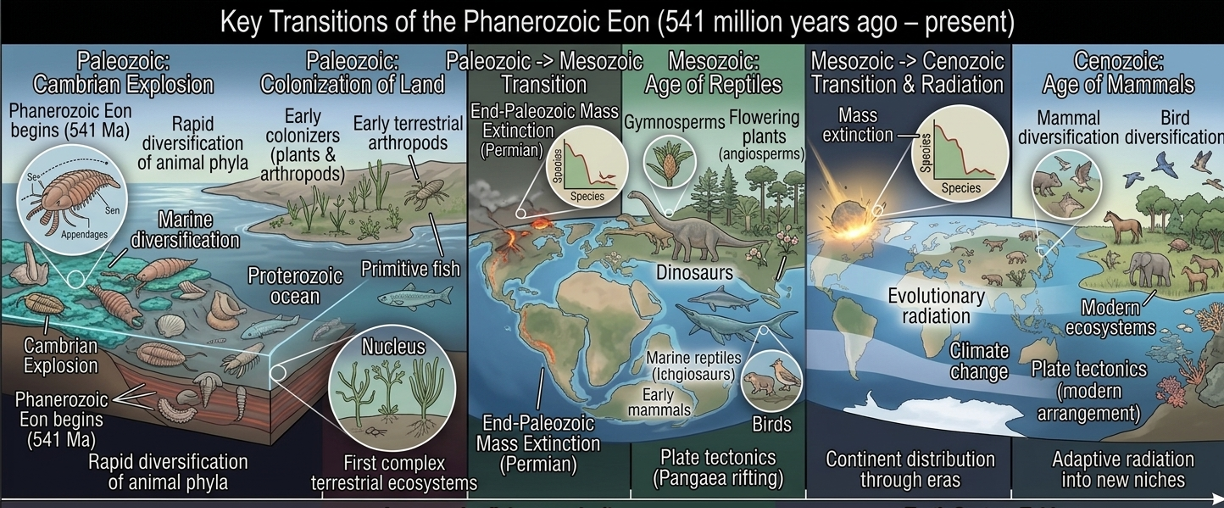
Figure 10. The Phanerozoic Eon. The Phanerozoic Eon is defined by the widespread presence and diversification of complex multicellular life. Across the Paleozoic, Mesozoic, and Cenozoic eras, life expanded from early marine diversification to dominant terrestrial ecosystems and finally to modern communities shaped by mammal and bird diversification. Throughout this eon, Earth systems including plate tectonics, climate change, and mass extinctions continually reshaped biodiversity, repeatedly driving evolutionary innovation and ecological restructuring.
The Phanerozoic Eon: The Paleozoic Era (541 – 252 Million Years Ago)
The Paleozoic Era (541 – 252 million years ago) marks the beginning of the Phanerozoic Eon and represents one of the most important periods of evolutionary expansion in Earth’s history. During this era, life rapidly diversified in the oceans, developed increasingly complex body structures, and eventually expanded onto land. The Paleozoic transformed Earth from a world dominated primarily by simple marine organisms into one containing diverse ecosystems of plants, invertebrates, fishes, amphibians, and early reptiles.
The Cambrian Explosion
The Paleozoic Era opened with the Cambrian Explosion, a relatively short interval of rapid evolutionary diversification in which nearly all major animal body plans appeared in the fossil record. During this time, organisms evolved complex features such as eyes, exoskeletons, segmented bodies, specialized appendages, and advanced forms of movement. Predation also became a major evolutionary force for the first time, fundamentally changing ecological interactions. As predators evolved new methods for capturing prey, prey organisms simultaneously developed defensive adaptations such as shells, armor, burrowing behaviors, and increased mobility. This evolutionary “arms race” accelerated natural selection and drove increasingly rapid diversification across marine ecosystems.
Fossil Evidence of Early Animal Life
Several exceptionally preserved fossil deposits from the early Paleozoic and late Proterozoic provide some of the clearest evidence for the emergence and diversification of complex animal life. These sites are especially important because most early organisms lacked hard shells or bones, meaning their soft tissues would normally decay before fossilization could occur. Under rare environmental conditions, however, rapid burial and low-oxygen environments allowed delicate anatomical structures to be preserved in remarkable detail, giving scientists an unprecedented view into early evolutionary history.
Read in chronological order, these fossil deposits show a clear progression in the early evolution of animal life, moving from simple multicellular beginnings to fully complex ecosystems. The earliest evidence comes from the Doushantuo Formation, which preserves microscopic fossils from the late Proterozoic just before the Cambrian Explosion. These include embryos, algae, and early multicellular structures that reveal life at a very early developmental stage. They show that key foundations of animal complexity (such as multicellularity, early cell specialization, and basic developmental patterns) were already present before large animals appeared in the fossil record.
Next, the Ediacara Hills preserve a group of large soft-bodied organisms that lived shortly before the Cambrian. These organisms represent a major step beyond microscopic life, showing that multicellular organisms had grown large enough to form visible ecosystems. However, their body structures are unusual and often do not match modern animal groups, suggesting they may represent early experiments in multicellular life or extinct evolutionary lineages. Even so, they show that ecosystems were already becoming more complex and capable of supporting larger organisms.
Finally, the Burgess Shale captures life during the middle Cambrian, when animal diversity expanded dramatically. This site preserves a wide range of soft-bodied organisms with highly developed features such as eyes, jointed limbs, segmented bodies, and specialized feeding structures. Unlike earlier deposits, the Burgess Shale shows fully established ecosystems with clear ecological roles, including predators, scavengers, filter feeders, and burrowers. The presence of active predators like Anomalocaris shows that strong ecological interactions were already driving evolutionary change.
Together, these fossil sites form a clear timeline. The Doushantuo Formation shows the early biological groundwork for complex life, the Ediacaran Biota shows the first large multicellular ecosystems, and the Burgess Shale reveals fully developed, diverse animal communities. Read in sequence, they show a gradual and continuous increase in biological complexity leading into the Cambrian Explosion.
The End of the Paleozoic: The Permian–Triassic Mass Extinction and a Global Reset of Life
The end of the Paleozoic Era, marking the transition into the Mesozoic, was defined by the most severe mass extinction event in Earth’s history: the Permian–Triassic extinction, often called “The Great Dying.” Around 252 million years ago, global ecosystems collapsed on an unprecedented scale, eliminating an estimated majority of marine species and a large fraction of terrestrial life. The causes were likely multiple and reinforcing, including massive volcanic eruptions in the Siberian Traps, rapid increases in greenhouse gases, ocean acidification, and widespread deoxygenation of the oceans.
As oxygen levels dropped in marine environments, complex ecosystems that had developed over hundreds of millions of years became increasingly unstable, leading to the collapse of food webs from the bottom up. Reef systems disappeared almost entirely, and many dominant Paleozoic groups such as trilobites and many types of brachiopods were driven to extinction. On land, the effects were equally devastating. Forests declined sharply, terrestrial vertebrate diversity collapsed, and ecosystems became simplified, with only a small number of resilient species surviving the environmental stress. The recovery that followed was slow and uneven, as surviving lineages gradually repopulated ecological niches left vacant by the extinction. However, this biological reset also created evolutionary opportunity: the ecological dominance of Paleozoic groups was permanently broken, allowing new lineages—particularly archosaurs and early relatives of mammals—to diversify in the early Mesozoic. In this way, the end of the Paleozoic represents both a catastrophic loss of biodiversity and a pivotal turning point that reshaped the trajectory of life on Earth.
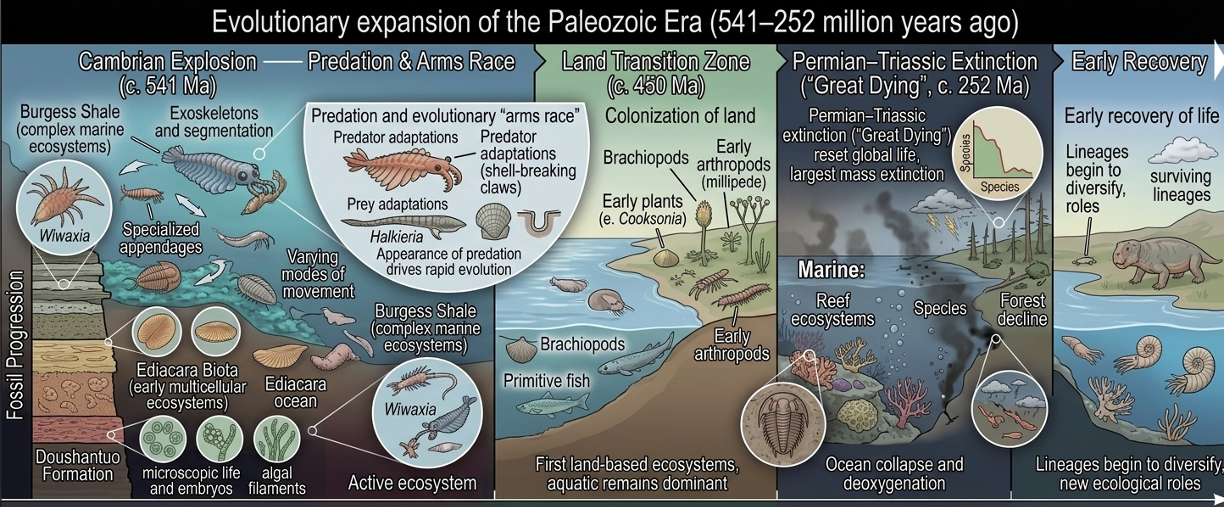
Figure 11. Paleozoic Era. The Paleozoic Era represents a major expansion of life, beginning with rapid marine diversification during the Cambrian Explosion and progressing through increasingly complex ecosystems and the colonization of land. Exceptional fossil sites such as the Doushantuo Formation, Ediacara Biota, and Burgess Shale document the stepwise increase in biological complexity. The era ends with the Permian–Triassic mass extinction, the most severe biodiversity loss in Earth’s history, which reshaped global ecosystems and opened ecological space for new groups in the Mesozoic Era.
The Phanerozoic Eon: The Mesozoic Era (252 – 66 Million Years Ago)
The Mesozoic Era (252 – 66 million years ago), often called the “Age of Reptiles,” represents a long interval of recovery, diversification, and ecological restructuring following the Permian–Triassic mass extinction. In its early stages, Earth’s ecosystems were still unstable and simplified, but over time life rebounded and expanded into new dominant groups. While dinosaurs eventually became the most iconic and ecologically dominant land animals, the Mesozoic was also a period of important evolutionary origins, including the first appearance of mammals, birds, and flowering plants. This era is traditionally divided into the Triassic, Jurassic, and Cretaceous Periods, each marked by distinct stages in the rise and transformation of global ecosystems.
Triassic Recovery and the Origins of Major Groups
The Triassic Period opened in the aftermath of the Permian extinction, when surviving organisms began rebuilding ecosystems from low diversity and high environmental stress. During this recovery phase, early dinosaurs, pterosaurs, and mammals emerged, alongside other surviving reptile lineages. Ecosystems were still relatively unbalanced, but these early groups established the evolutionary foundations for later dominance. Archosaurs, in particular, diversified rapidly and set the stage for the rise of dinosaurs as major terrestrial vertebrates. Mammals also originated during this time, though they remained small and ecologically limited compared to dominant reptile groups.
Jurassic Expansion and Dinosaur Dominance
The Jurassic Period marked the height of dinosaur dominance and the development of more stable, complex ecosystems. Dinosaurs diversified into a wide range of herbivorous and carnivorous forms, occupying nearly all major terrestrial niches. Large sauropods dominated many landscapes, while theropods became highly successful predators. On land, lush forests expanded, supported largely by gymnosperm plants such as conifers, cycads, and ginkgos. In the oceans, marine reptiles such as ichthyosaurs and plesiosaurs became dominant predators, while pterosaurs ruled the skies as the first large flying vertebrates. This period reflects a fully established Mesozoic world in which reptiles were the primary drivers of ecological structure across land, sea, and air.
Cretaceous Innovation and Ecosystem Transformation
The Cretaceous Period brought further evolutionary innovation and ecological complexity. Flowering plants (angiosperms) appeared and rapidly diversified, fundamentally transforming terrestrial ecosystems. Their new reproductive strategies involving flowers and fruit increased pollination efficiency and seed dispersal, driving co-evolution with insects such as bees, butterflies, and beetles. At the same time, mammals and birds diversified, expanding into a wider range of ecological roles while still living in ecosystems dominated by dinosaurs. By the Late Cretaceous, global ecosystems had become highly complex, with intricate food webs spanning land and sea and strong interactions between plants, herbivores, and predators.
The End of the Mesozoic
The Mesozoic Era ended abruptly with a mass extinction event around 66 million years ago, marking one of the most significant turning points in Earth’s history. This extinction is widely linked to a massive asteroid impact at the Chicxulub site, combined with intense volcanic activity in the Deccan Traps. These events triggered rapid climate disruption, including blocked sunlight, global cooling, and ecosystem collapse. Non-avian dinosaurs, along with many marine and terrestrial groups, went extinct. However, some lineages—especially mammals, birds, and surviving reptiles—persisted, setting the stage for the next major chapter in Earth’s history: the rise of modern ecosystems in the Cenozoic Era.

Figure 12. The Mesozoic Era. The Mesozoic Era is defined by recovery from the Permian extinction, the rise and dominance of dinosaurs, and major evolutionary innovations including mammals, birds, and flowering plants. Across the Triassic, Jurassic, and Cretaceous periods, ecosystems became increasingly complex and interconnected. The era ended with a mass extinction event around 66 million years ago that eliminated non-avian dinosaurs and many other groups, reshaping global ecosystems and enabling the rise of modern life in the Cenozoic Era.
The Phanerozoic Eon: The Cenozoic Era (66 Million Years Ago – Present)
The Cenozoic Era (66 million years ago to the present) marks the most recent chapter of Earth’s history and is often called the “Age of Mammals.” It began in the aftermath of the end-Cretaceous mass extinction, when the disappearance of non-avian dinosaurs opened vast ecological niches across land, sea, and air. In this newly reorganized world, mammals and birds rapidly diversified, expanding into environments and ecological roles that had previously been dominated by reptiles. Over time, Earth’s climate gradually cooled and became more variable, shaping evolutionary pathways and driving the development of modern ecosystems.
Paleogene Expansion of Mammals
The Paleogene Period was defined by rapid mammalian diversification following the extinction of the dinosaurs. With reduced competition, mammals expanded in size, complexity, and ecological range, occupying niches as large herbivores, apex predators, tree-dwellers, and even fully aquatic forms such as early whales. Birds also diversified significantly, becoming major components of terrestrial and aquatic ecosystems. Forested environments dominated much of the planet, supporting rich and rapidly evolving ecosystems in which modern mammal groups began to take shape.
Neogene Ecosystem Restructuring and Grasslands
The Neogene Period was marked by important climatic cooling and the spread of grasslands across many regions of the world. This shift in vegetation had major evolutionary consequences, particularly for herbivorous mammals, which adapted to grazing lifestyles with specialized teeth, digestive systems, and social behaviors. These open habitats also favored fast-moving predators and drove new forms of ecological interaction. During this time, early hominins appeared in Africa, representing the beginning of the human evolutionary lineage within a broader context of primate diversification.
The Quaternary and the Rise of Humans
The Quaternary Period is characterized by repeated ice ages and rapid environmental fluctuations that shaped modern ecosystems. Glacial cycles caused frequent shifts in sea levels, vegetation zones, and species distributions, placing strong selective pressures on many organisms. It is within this dynamic and unstable environment that Homo sapiens emerged, evolving advanced cognitive abilities, tool use, and complex social structures. Modern humans eventually spread across the globe, adapting to a wide range of environments and beginning to exert large-scale influence on ecosystems, marking the start of a new phase in Earth’s history defined by both biological and environmental transformation.

Figure 13. The Cenozoic Era. The Cenozoic Era represents the modern chapter of Earth’s history, beginning with the recovery of ecosystems after the extinction of non-avian dinosaurs. Mammals and birds diversified into a wide range of ecological roles, while global climate cooling led to the expansion of grasslands and the restructuring of terrestrial ecosystems. Repeated ice age cycles during the Quaternary shaped modern biodiversity and environments. Within this dynamic context, Homo sapiens emerged and spread globally, increasingly influencing ecosystems and marking a new phase of biological and environmental change.