
Chapter 4: Mendel and the gene

- Lecture Video: Mendel and the gene
- Lecture Slides: Mendel and the gene
- Study Guide: Mendel and the gene
- Lab: Mendel and the gene
The “gene”
The word gene originates from ancient Greek, meaning locus, or region. The concept of the gene originated with the work of Gregor Mendel in his discoveries of inheritance patterns. While he never used the term, gene, he developed the basis of its modern context. While looking at the inheritance of a variety of physical characteristics of pea plants (i.e. flower color, seed shape, etc), he discovered that the inheritance was best explained by the presence of “discrete, heritable units.”
The widely accepted inheritance hypothesis of the day was known as blending inheritance, which predicted that the traits of parents blended in the offspring. For example, a white flowered plant and a purple flowered plant would produce an intermediate, or light purple flowered plant. To test this hypothesis, Mendel carried out a series of experiments on the common pea plant in his monastery’s experimental garden plot between 1856 and 1863.
Pure lines
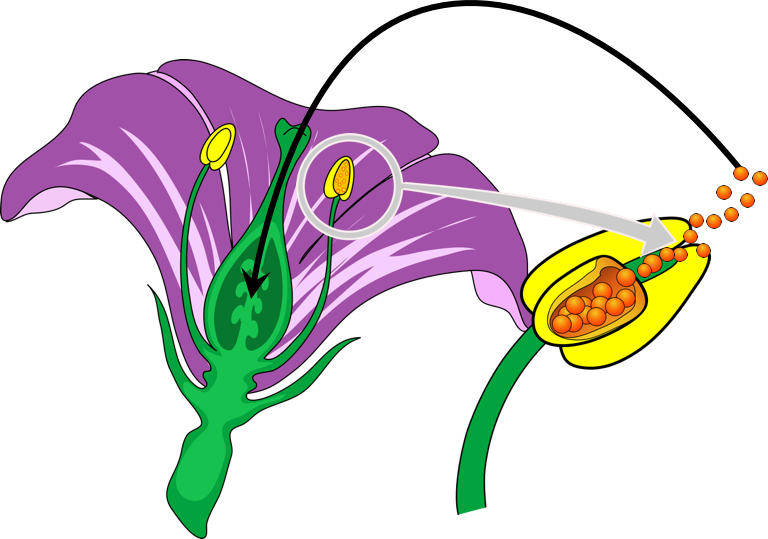
Figure 1. Self-pollination produces pure lines. Mendel completed a controlled self-pollination to produce true-breeding plants, which exhibited identical phenotypes (or physical characteristics) as the parent.
The pea plant awarded Mendel several advantages for studying inheritance. First, pea plants grow easily and quickly allowing for many replicates and generations in a relatively short amount of time. Second, peas have many different physical characteristics, or phenotypes. Mendel eventually analyzed seven phenotypes: seed shape, seed color, flower color, flower location, pod shape, pod color, and plant height. Most importantly, he could obtain true-breeding varieties, or pure lines. When these seeds were planted, they produced known phenotypes. For example, pure line seeds for purple flowers would always develop into plants with purple flowers. Pure lines are produced by self-pollination, where a flower is covered and the pollen of a plant is used to fertilize the same plant, essentially producing a clone. Therefore, whatever phenotypes the self-fertilized plant had, the seeds would produce plants with exactly the same physical characteristics. These true-breeding pure lines are known as the parent generation, or P generation.
Mendel initially focused his analysis on seed shape. Pea seeds are either round (smooth) or have angular indentions (wrinkly). After self-pollination, he grew out pure line seeds of both varieties until they flowered. To control against unwanted pollination, the developing flowers were covered with mesh bags. Once fertile, pollen was collected with a sterilized paint brush from the flowers of one phenotype (i.e. smooth seeds) and used to pollinate the opposite phenotype (i.e. wrinkly seeds), producing a hybrid of the pure lines. He also cross pollinated smooth seeded varieties with wrinkly seeded varieties too. Once pollinated, the mesh bags were reinstalled and the seeds allowed to develop. These hybrids are notated as the first filial generation, or the F1 generation.

Figure 2. Mendel's results did not support blending inheritance. Rather than getting intermediate phenotypes in the F1 generation, Mendel discovered that one of the original phenotypes from the P generation was present. This discovery led to his Principle of Dominance.
F1: hybrids of pure lines
While it was known that crossing similar true-breeding plants would produce the same phenotypes, or pure lines, the blending inheritance hypothesis predicted that crossing two different pure lines would produce an intermediate in the F1 generation. Mendel expected to get an intermediate result of slightly wrinkly seeded plants from a cross of pure line smooth seeded plants with pure line wrinkly seeded plants. Mendel’s results conflicted with this prediction. Once the seeds developed from Mendel’s monohybrid cross, he discovered that all the resultant seeds (the hybrids) were not semi-wrinkly. Rather, they were identical in shape to the round seeded pure lines. From his controlled pollination, he knew the hybrids had equal components of the wrinkly seeded and smooth seeded parent pure lines. He concluded the smooth seed phenotype masked the expression of the wrinkly seed phenotype. He termed the expressed phenotype of the hybrids (smooth seeds in this case) as the dominant phenotype (notated as a capital letter, e.g. A), whereas the repressed phenotype (wrinkly seeds), was termed the recessive phenotype (notated as a lower-case letter, e.g. a). This result directly contradicted the blending inheritance hypothesis. Hybrids were not intermediates.
The Principle of Dominance

Figure 3. Principle of Dominance. A phenotype of an individual is determined by the combination of alleles, known as genotype. During fertilization, gametes carry exactly one allele from each parent. If both alleles are the same (homozygous), that specific phenotype is expressed. If an organism has one of each allele (heterozygous), the organism expresses the dominant characteristic.
From this initial study, Mendel concluded that the physical characteristics (or phenotypes) he was studying (i.e. seed shape, flower color, etc.), were determined by the combination of what he referred to as “particles” which he defined as “discrete heritable units.” This concept, known as the particulate inheritance hypothesis, was in direct contrast to blending inheritance, which predicted that phenotypes of offspring are intermediate to the parents. We now refer to Mendel’s particles as alleles, alternate versions of a gene. Mendel discovered the presence of dominant and recessive alleles. Furthermore, he predicted that an offspring receives exactly one allele from each parent, giving each individual two alleles. The combination of those alleles is known as an individual’s genotype, of which there are three possibilities: two dominant alleles termed homozygous dominant (notated as with two capital letters, e.g. AA), or two recessive alleles termed homozygous recessive (e.g. aa), or one of each allele termed heterozygous (e.g. Aa). From his monohybrid cross, Mendel predicted that both the homozygous dominant (e.g. AA) and the heterozygous (e.g. Aa) genotypes would produce the dominant phenotype (e.g. round seeds), while the recessive phenotype (e.g. wrinkled seeds) is only expressed by the homozygous recessive genotype (e.g. aa). This became known as Mendel’s Principle of Dominance, where a genotype is composed of two alleles and if the alleles differ only one phenotype will be expressed (dominant) while the other is masked (recessive). Blended inheritance predicts the heterozygote (e.g. Aa) would be an intermediate between the two homozygous phenotypes (e.g. slightly wrinkled seeds).
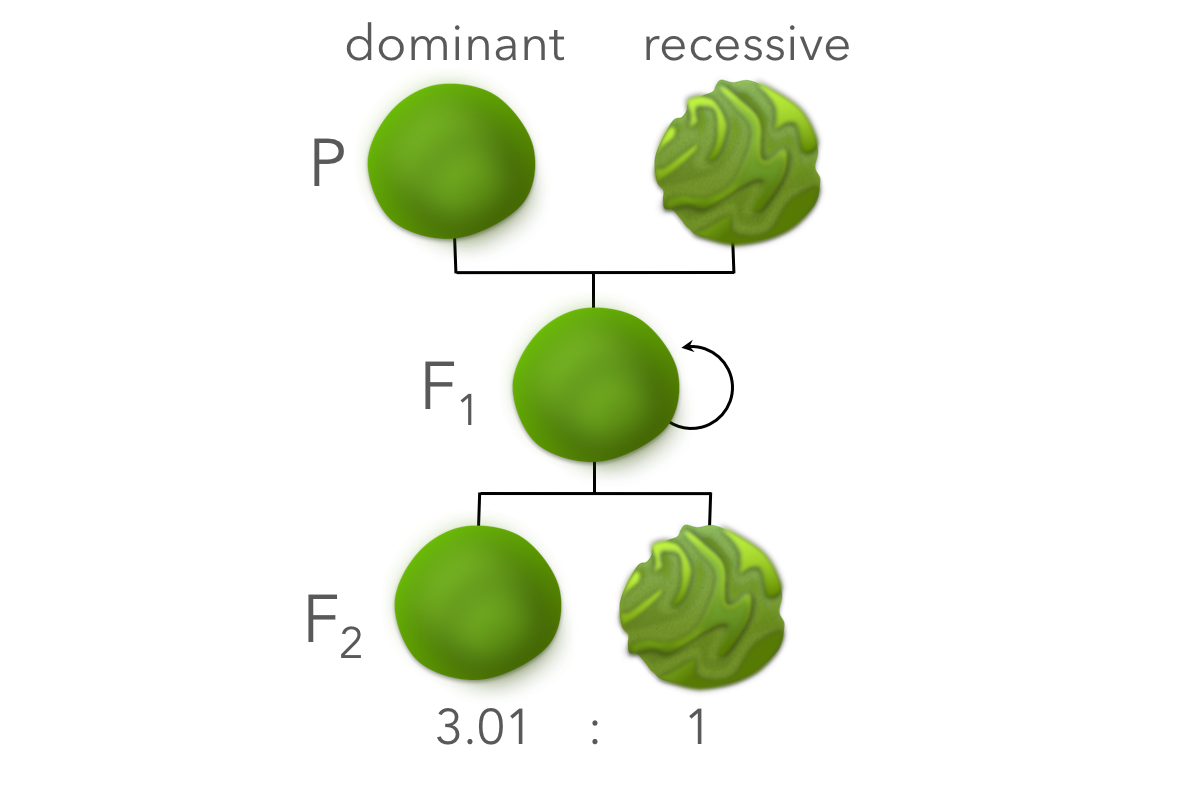
Figure 4. Phenotypic expressions of Mendel's experiment. Seeds resultant from self-pollination were grown out producing pure lines of the P generation. Cross-pollination of the P generation produced the F1 generation (hybrids) which expressed round seeds, indicating round as the dominant phenotype and wrinkly as the recessive phenotype. F1 plants were crossed producing the F2 generation, in which approximately 3 dominant phenotypes were expressed for every recessive.
F2: hybrids of hybrids
After this discovery, Mendel grew out the F1 generation (the hybrids of the pure lines) and then repeated the same cross pollination, generating new seeds. These seeds formed the second filial or F2 generation, which can be thought of as hybrids of the original hybrids. To his surprise, not only did the dominant genotype (round seeds) emerge in the F2 generation, but also the recessive phenotype (wrinkly seeds). More precisely, he collected 5474 round seeds and 1850 wrinkly seeds, or a 2.96 dominant to 1 recessive ratio, approximately 3:1.
Mendel then repeated the same experiment for a variety of other traits with his peas, such as: seed color, pod color, flower color, and pod shape. From his original experiment, he predicted that in each of the physical characteristics one phenotype would dominate in the F1 generation, masking the recessive phenotype. From his seed shape experiment, he also predicted an approximate 3:1 ratio of dominant to recessive phenotypes in the F2 generation. And that is exactly what he found. Let’s use seed color as an example. Mendel crossed pure line yellow seeded and green seeded plants. Seeds from that cross (the F1 generation) were all yellow, indicating the dominant phenotype. After growing those seeds out and crossing the hybrids, he counted the F2 seeds: 6022 yellow and 2001 green, a 3.01:1 ratio. He found a similar ratio for other characteristics in the F2 generation: pod shape was 2.95 inflated to 1 constricted, pod color was 2.82 green to 1 yellow, and flower color was 3.15 purple to 1 white – all approximately 3:1.
The Punnett Square
Mendel’s discoveries went largely discounted and forgotten, due to the strong acceptance of a competing inheritance hypotheses - more on that later. At the turn of the 20th century, researchers designed experiments that led them to the same conclusions Mendel arose at decades earlier, without knowing of Mendel’s work (we think). Eventually his publications were rediscovered by geneticists and he was given due credit. One of those scientists was Reginald Punnett, who developed a simple tool, now known as the Punnett Square, to predict the probability of genotypes and phenotypes from controlled crosses.

Figure 5. Punnett square of P generation cross. FF X ff produces the F2 generation.
Let’s revisit Mendel’s monohybrid cross of flower color, using a Punnett square to visualize Mendel’s predictions. From the results of his F1 generation, Mendel determined that the purple phenotype for flower color was dominant, while white flower color was recessive. Based on his Principle of Dominance, purple flowers could be either the homozygous dominant (FF) or heterozygous (Ff), and white flowers were homozygous recessive (ff).
When Mendel self-pollinated the pea plants to produce a pure line for the P generation, he was creating a homozygote, either FF or ff depending on the original plant. Once the pure lines reached reproductive maturity, he cross pollinated a homozygote dominant with a homozygote recessive, notated as FF x ff. Once the seeds matured and were allowed to develop into plants (the F1 generation), they all expressed the purple flower phenotype, but were known to be hybrids (Ff). He concluded that one of the dominant alleles (F) from the homozygous dominant parent (FF) and one of the recessive alleles (f) from the homozygous recessive parent (ff) were both present in the plants of the F1 generation. In other words, all plants in the F1 generation were heterozygotes (Ff), and expressed the dominant phenotype for flower color: purple.

Figure 6. Punnett square of a monohybrid cross. Ff X Ff produces the F2 generation.
Next, he crossed the plants of the F1 generation: Ff x Ff, producing offspring that developed into the F2 generation. To calculate predictions of this cross we use a Punnett Square. Mendel predicted that there was an equal probability of an offspring receiving either allele present in the parent. If a parent was heterozygous (i.e. Ff), each offspring had a 50% chance of getting a dominant allele (F) and a 50% chance of receiving a recessive allele (f). In the Punnett Square this is visualized by separating the alleles of one parent and placing them above a box bisected vertically. If the other parent is also heterozygous, the box is also bisected horizontally, and the alleles of the other parent are placed one on top of another to the left of the box, representing the equal probability of receiving that parent’s alleles. The four boxes in the middle represent the probability the offspring will acquire a specific genotype. Once the parental alleles are in place, transcribe the top alleles down into each internal box below it and the side alleles to both boxes to its right. In this case, we end up with one box that is homozygous dominant (FF), two boxes that are heterozygous (Ff) and one box that is homozygous recessive (ff). With this tool, we can predict the probable genotypic ratio of the offspring: 25% FF to 50% Ff to 25% ff (simplified as 1 FF: 2 Ff: 1 ff). However, Mendel could not see the alleles but could detect the phenotypes. Since both FF and Ff appear purple and ff appears white, he could predict the phenotypic ratio: 75% purple-flowered plants to 25% white-flowered plants (3 purple: 1 white). This explains the approximate 3:1 ratio he detected among several different phenotypes.

Figure 7. Principle of Segregation. Mendel discovered organisms have two copies of a gene (but potentially different alleles, as is the case with heterozygotes (Aa). In the production of gametes, each gamete receives exactly one copy of a gene, at random. During fertilization, gametes fuse at random producing a new organism. The Principle of Segregation explains Mendel's 3 dominant: 1 recessive ratio in the F2 generation.
The Principle of Segregation
The fact that Mendel observed this consistent 3:1 phenotypic ratio in the F2 generation across several different traits allowed him to expand the particulate inheritance model. In what has become known as Mendel’s Principle of Segregation, Mendel predicted that an organism’s physical traits are passed from the parental generations as ‘discrete particles’ (now referred to as alleles), with exactly one particle from each parent in each gamete (i.e. pollen or egg cells). Therefore, each gamete has an equal probability (50%) of receiving either allele present in the parent. When an organism generates a gamete only one of the two possible parental alleles is allocated to each gamete. Furthermore, the allele allocation is random, which explains why there is an equal probability of receiving either allele. When gametes fertilize a new organism is formed, which have genotypes that are the combination of parental alleles.
Dependent vs. Independent Assortment: the dihybrid cross

Figure 8. Prediction of dependent assortment. In a dihybrid cross (FfSs X FfSs), the dependent assortment predicts the F and the S alleles are physically linked, and the f and the s alleles are linked. When a dihybrid cross is conducted, dependent assortment predicts offspring with a phenotypic ratio of 3 purple-flower/yellow-seed: 1 white-flower/green-seed.
After Mendel discovered the principles of segregation and dominance using his monohybrid crosses, the next big question was whether or not the inheritance of multiple phenotypes are linked. There were two alternative hypotheses: dependent and independent assortment. The dependent assortment hypothesis predicts that the inheritance of physical characteristics are linked during gamete formation. For example, an allele that codes for a specific flower color is connected to an allele for seed color. Alternatively, independent assortment predicts alleles for different phenotypes are sorted into gametes independently from one another. In other words, receiving an allele for one phenotype (e.g. flower color) does not determine the presence of another phenotype (e.g. seed color).

Figure 9. Prediction of independent assortment. Independent assortment predicts the alleles are not physically linked. All combinations are possible: FS, Fs, fS, and fs. When a dihybrid cross is conducted, independent assortment predicts offspring with a phenotypic ratio of 9 purple-flower/yellow-seed: 3 purple-flower/green-seed: 9 white-flower/yellow-seed: 1 white-flower/green-seed.
To test these competing hypotheses Mendel conducted a dihybrid cross. He identified pea plants that had dominant phenotypes for two characteristics and self-pollinated them. The plants resultant from those seeds were pure lines for two dominant characteristics. In this case the pure line pea plants of the P generation would be homozygous dominant for purple flowers (FF) and homozygous dominant for yellow seeds (SS). These plants are notated FFSS. The same procedure was conducted with plants that had recessive phenotypes for both characteristics, producing pure lines with white flowers and green seeds: ffss.
Next the P generation was crossed and grown out producing the F1 generation of dihybrids: all FfSs. As expected all of the dihybrids expressed the dominant phenotypes: purple flowers and yellow seeds. When the F1 generation reached maturity, he conducted a controlled cross of the dihybrids and grew them out, producing the F2 generation.
The competing alternative hypotheses predicted different results for the F2 generation. If alleles were linked (as predicted by dependent assortment), the F allele would be physically connected to the S allele in the production of a gamete, and the f allele and s allele would be linked in another gamete. Therefore, in a dihybrid cross the pollen and eggs would either have allele combination F and S (FS) or f and s (fs). No other combinations are possible: neither Fs nor fS. If dependent assortment was supported by this experimental design, the predicted genotypic ratio would be 1FFSS: 2FfSs: 1ffss and the phenotypic ratio (what he could actually observe) would be 3 purple flower/yellow seeded plants to 1 white flower/green seeded plants, which can be generalized as 3 DD (dominant/dominant): 1 RR (recessive/recessive) for other phenotypic combinations under investigation. Dependent assortment would not produce plants with a combination of dominant and recessive phenotypes from a dihybrid cross. In other words, there would be no purple flowered, green seeded plants nor would there be white flowered, yellow seeded plants.
In contrast, the independent assortment hypothesis predicted that all allele combinations were possible in gamete production: FS, Fs, fS, and fs. This expands the Punnett square from four boxes to 16. The prediction of the phenotypic ratio of independent assortment in this case would be 9 purple/yellow: 3 white/yellow: 3 purple/green: 1 white/green (generalized as 9DD: 3DR: 3RD: 1RR). Results of his dihybrid cross were consistent with independent assortment, indicating that alleles for different phenotypes segregate independently of each other. This hypothesis became known as Mendel’s Principle of Independent Assortment.
Particulate Inheritance: a summary
Mendel’s elegant experiments clearly disproved the blending inheritance hypothesis. He proposed an alternate hypothesis: the particulate inheritance hypothesis, which has three main components:
- The Principle of Dominance predicts that each phenotype is determined by two alleles, one of which is expressed when present (known as the dominant allele). Recessive phenotypes are expressed only when an organism is homozygous recessive.
- The Principle of Segregation predicts that during gamete production, each gamete receives exactly one of the two alleles present in the parental cell at random.
- The Principle of Independent Assortment predicts alleles coding for different phenotypes are segregated independently from one another during gamete formation.
Pedigree Analysis
Mendel’s success was in part due to the pea plants’ ability to self-pollinate, creating pure lines. However, nearly all animals are incapable of self-fertilization. This makes understanding the genetics of animals (like humans) increasingly more complicated than plants.

Figure 10. Pedigree analysis. Understanding Mendelians genetics is a useful tool for deconstructing genotypes and phenotypes in a family history.
Pedigree analysis is a method that uses the principles developed by Mendel to look at families’ genetic histories with an attempt to understand how phenotypes are expressed. In a pedigree, squares represent males and circles represent females. Horizontal lines represent a mating event producing children. Children are connected to their parents’ horizontal line by a vertical line and are shown below their parents. Generations are shown with Roman numerals (i.e. I, II, III, etc.). Pedigree analyses are typically used to follow a specific genetically inherited trait, which is customarily shaded.
If two parents have the same phenotype (e.g. Fig. 10; 1 and 2) and produce a child that has a different phenotype (Fig. 10; 6 and 8), we know that the child’s phenotype must be the recessive expression. Furthermore, we know that the child’s genotype must be homozygous recessive (i.e. aa), and the genotypes of both parents must be heterozygous (i.e. Aa). Using this logic, we can begin to deconstruct larger pedigrees and predict its members’ genotypes, based on their relationships with individuals containing the recessive phenotype. In this example, all of the members with the genetically inherited trait are homozygous recessive (Fig. 10; 4, 6, 8, and 11). Each individual that is homozygous recessive received exactly one recessive allele from each parent. Therefore, if a child has a recessive phenotype and both parents have a dominant phenotype, those parents must have been heterozygous (Fig. 10; 1 and 2). If we know that the parents are heterozygous, we cannot determine the exact genotype of a child that expresses the dominant phenotype without further information (Fig. 10; 5 and 7). The child could either be homozygous dominant or heterozygous. His genotype is denoted A?. If the parents have both the recessive (Fig 10; 8) and dominant phenotype (Fig. 10; 9), and their child has a dominant phenotype (Fig. 10; 9, 10, and 12), we know that child’s genotype is heterozygous because one recessive allele must have come from the parent with the recessive phenotype and the dominant allele from the parent with the dominant phenotype. Pedigree analysis is a useful tool in genetic counseling and understanding human inheritance.
Mendel’s discoveries were discounted

Figure 11. Darwin's pangenesis inheritance hypothesis. Darwin believed gametes received 'gemmules' from each organ of the body. This belief incorporated concepts of blending inheritance and Lamarck's inheritance of acquired characteristics, which were contradictory to Mendel's particulate hypothesis.
Mendel’s discoveries went largely unnoticed or ignored, due to the strong acceptance of the competing inheritance hypotheses within the scientific community: blending inheritance . Acceptance of the particulate inheritance model was also very likely discounted due to his ignoble status among the academic elite of the day. At the time of Mendel’s publication (in 1866), the major topic of conversation in the biological community was Charles Darwin’s theory of natural selection as described in On the Origin of Species (published in 1859). While the theory of natural selection was gaining greater acceptance, Darwin published his own theory of inheritance known as pangenesis (1868). In his description, he suggested that all organs in the body contain unseen ‘gemmules’ that are extracted and travel to the reproductive cells. In this manner, a reproductive cell would consist of many ‘gemmules’ from all organs and tissues of the body. In humans, for example, sperm and egg cells would acquire particles from all the parts of the body (the heart, skin, feet, etc.) and during fertilization, these ‘gemmules’ would mix forming a new organism.
Darwin’s pangenesis theory was an extension of his grandfather’s beliefs. In 1794 Erasmus Darwin (in Zoonomia) suggested that “animals have arisen from one living filament…with power of acquiring new parts” based on the animals’ interactions with its environment and each acquired acquisition is inherited by successive generations. In his widely acclaimed book, Philosophie Zoologique (in 1809), Jean-Baptiste de Larmarck expanded this concept, creating the first formal theory of inheritance: the inheritance of acquired characteristics. Essentially, Lamarck suggested that individuals acquire physical characteristics during their lifetime through their interactions with the environment, and those acquired characteristics were inherited by their offspring. The giraffe’s neck is a classic example. Lamarck argued that giraffes stretch their neck to reach higher food sources, eventually leading to a longer necked individual. He posited that this acquired characteristic was passed to the next generation, a trend which continued in successive generations eventually leading to the long-necked giraffes we see today.
Mendel’s study was diametrically opposed Darwin’s pangenesis theory which supported both the inheritance of acquired characteristics and blended inheritance, even though Darwin had no experimental analyses to support these concepts. Furthermore, Mendel rejected Darwin’s concept of natural selection, arguing that species are fixed and do not change over time. While Mendel’s work was known about by his academic contemporaries, his conclusions were essentially neglected as a result of a lowly monk’s clear opposition to the biological superstar of the day, Charles Darwin. After several decades, Mendel’s laws were rediscovered by Walter Sutton and Theodor Boveri in 1900 leading to a widespread rejection of Darwin’s pangenesis theory. The rediscovery of Mendelian genetics combined with advances in microscopy and experimental design allowed Sutton and Boveri to establish a more accurate genetic model, known as the chromosome model of inheritance. This is the topic of our next chapter.
© 2018. www.thebiologyprimer.com. All rights reserved.