
Chapter: The Cell Cycle
Binary Fission in Prokaryotes
Prokaryotic (i.e. bacteria) cell division, which occurs via binary fission. Binary fission is an asexual reproductive process characteristic of Bacteria and Archaea, and it reflects a fundamentally simpler organizational framework due to the absence of a nucleus and membrane-bound organelles. In binary fission, the single circular chromosome is replicated from a defined origin of replication. The two resulting DNA molecules are actively segregated as the cell elongates, with partitioning mechanisms ensuring their movement toward opposite regions of the cytoplasm. Concurrently, the plasma membrane expands, and ultimately invaginates along with the cell wall to produce two genetically identical daughter cells. Although structurally simpler than mitosis, binary fission is highly efficient and produces clonal populations. Genetic variation in prokaryotes does not arise from the division process itself but rather from replication errors (mutations) and horizontal gene transfer mechanisms such as transformation, transduction, and conjugation.
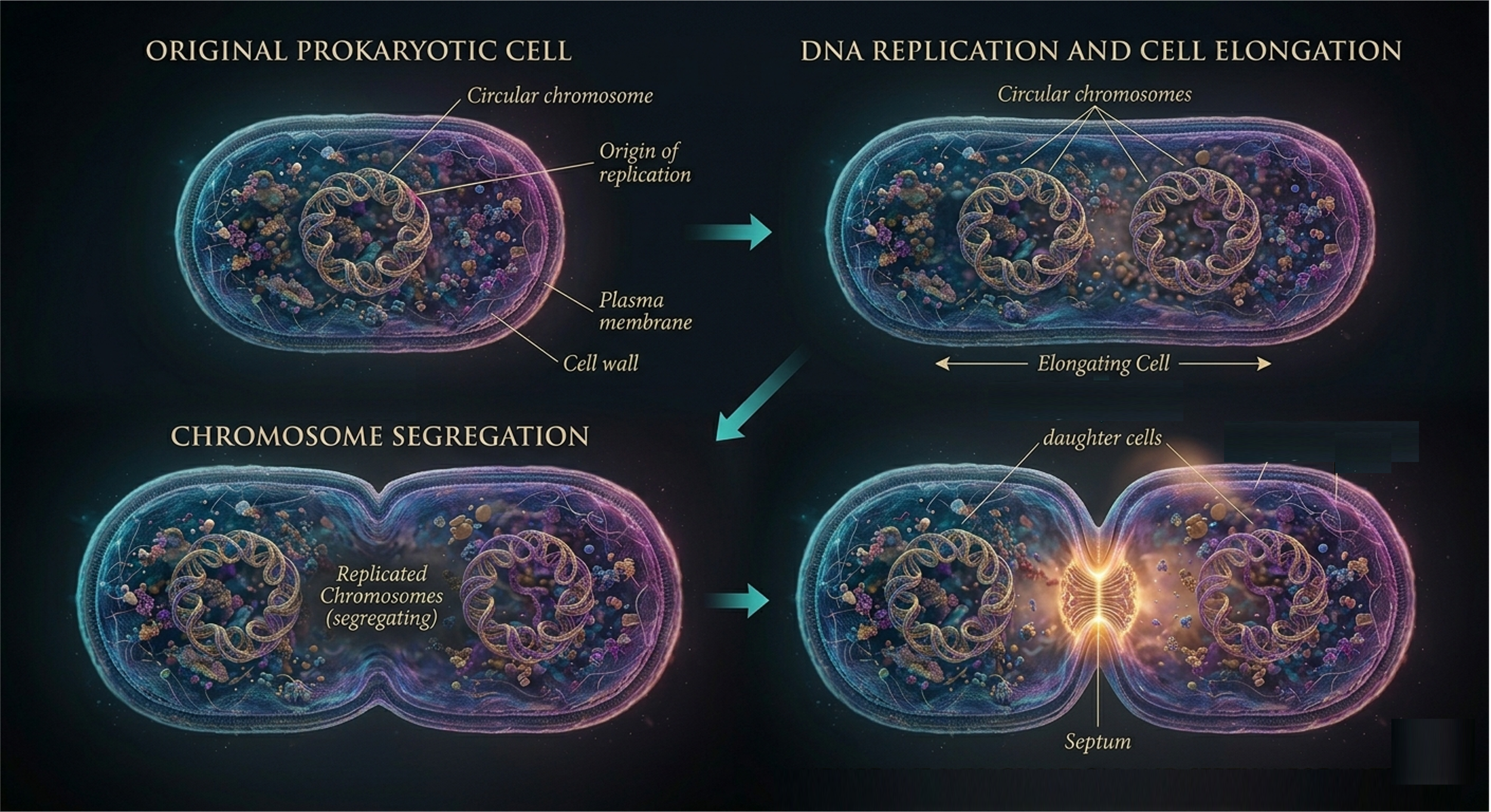
Figure 1. Binary Fission. Binary fission in prokaryotes involves replication of a single circular chromosome, segregation of DNA as the cell elongates, and division of the plasma membrane and cell wall to produce two genetically identical daughter cells. Although structurally simpler than mitosis, binary fission efficiently generates clonal populations, with genetic variation arising primarily through mutation and horizontal gene transfer.
Eukaryotic Cell Division: Overview
In eukaryotic cell division one parent cell produces two genetically identical daughter cells. This process is important because each daughter cell receives a complete and accurate copy of the DNA, helping maintain normal tissue structure and function. In unicellular eukaryotes, mitosis serves as a form of asexual reproduction. In multicellular organisms, it is primarily responsible for growth, development, tissue repair, and cell replacement.
In multicellular organisms, a single fertilized egg divides repeatedly through mitosis to produce the trillions of cells that make up a complex organism. These cells later specialize into different cell types through differentiation. In adults, mitosis continues to replace old, damaged, or worn-out cells. Some tissues, such as skin and blood, are constantly renewed, while others, such as many neurons, divide very little. Bone tissue also undergoes continuous remodeling through the coordinated activity of specialized cells. Together, these processes keep multicellular organisms functioning and maintaining tissue homeostasis.
Mitosis differs from meiosis, which is used for sexual reproduction. Mitosis maintains the chromosome number and produces genetically identical somatic cells, whereas meiosis reduces the chromosome number by half to produce haploid gametes such as sperm and egg cells. Fertilization then restores the diploid chromosome number and begins a new organismal life cycle.
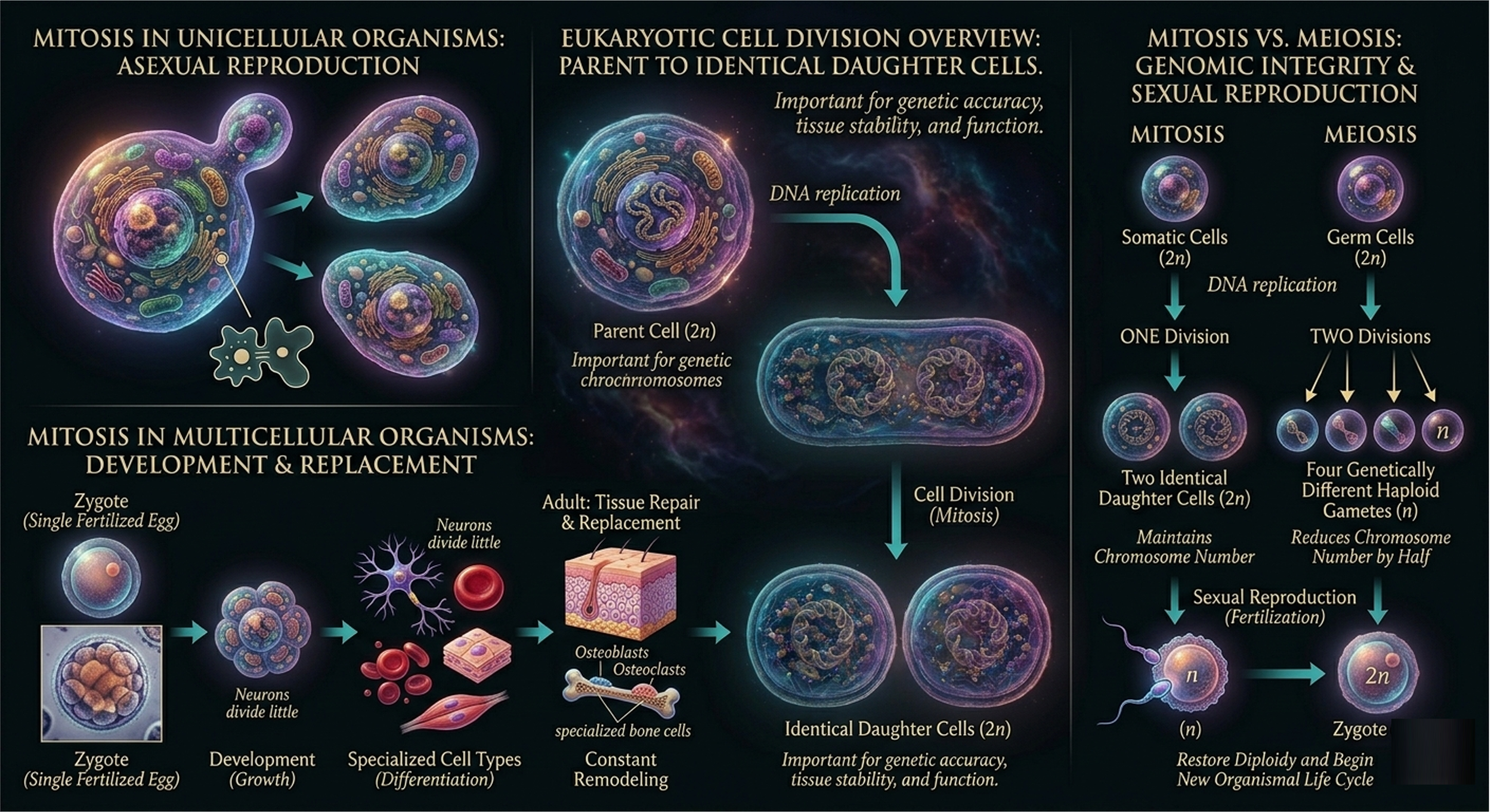
Figure 2. Eukaryotic cell division. In eukaryotes, cell division consists of interphase, mitosis, and cytokinesis. DNA is replicated before mitosis, and duplicated chromosomes are equally distributed into two genetically identical daughter cells.
The Cell Cycle
Eukaryotic cell division is embedded within a highly regulated sequence of events known as the cell cycle. The cell cycle ensures fidelity of DNA replication and accurate segregation of genetic material. It is conventionally divided into three major phases: interphase, mitosis (M phase), and cytokinesis. Interphase represents the longest portion of the cycle and encompasses cellular growth, metabolic activity, and preparation for division. Mitosis is the nuclear division phase in which duplicated chromosomes are segregated into two genetically identical nuclei. Cytokinesis follows mitosis and completes the division process by physically partitioning the cytoplasm into two daughter cells.
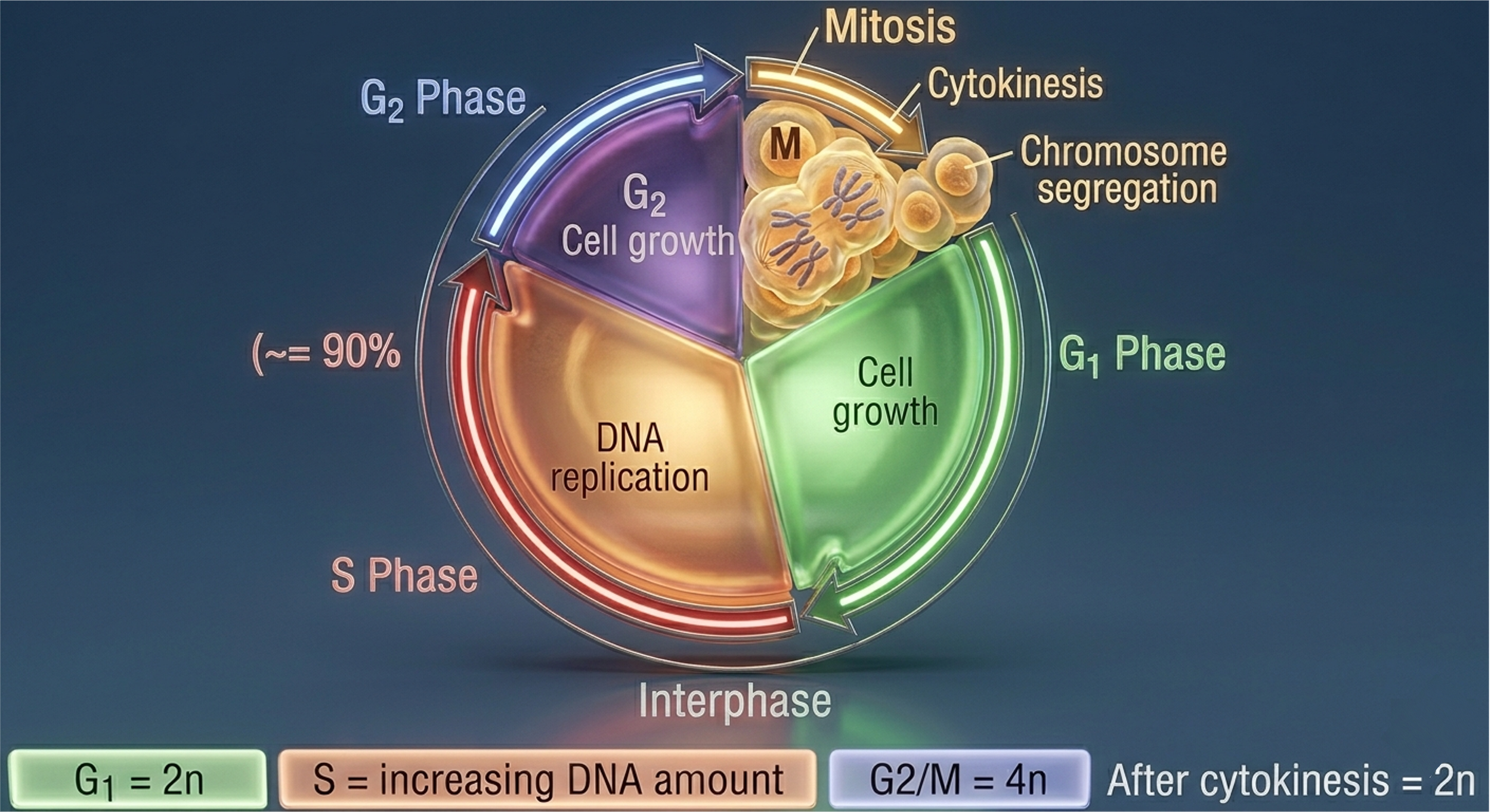
Figure 3. The eukaryotic cell cycle. Interphase includes G1, S, and G2 phases, during which the cell grows and replicates its genome. Mitosis segregates duplicated chromosomes, and cytokinesis physically divides the cytoplasm to produce two daughter cells.
Interphase: Cellular Growth and Genome Replication
Interphase is not a quiescent state but rather a period of intense biochemical activity. During interphase, cells carry out specialized metabolic functions, respond to extracellular signals, and coordinate the replication of their genome. Chromatin remains in a decondensed state, allowing transcriptional accessibility and DNA replication machinery to operate efficiently. Interphase is subdivided into three sequential phases: G1, S, and G2. Each phase represents a distinct functional stage of growth, genome duplication, and preparation for mitosis.
G1 Phase (Gap 1)
The G1 phase immediately follows mitotic division and represents a period of cellular growth and metabolic activity. During G1, the cell increases in size, synthesizes RNA and proteins, and produces enzymes required for DNA replication. Importantly, G1 also functions as a regulatory checkpoint in which the cell assesses environmental conditions, nutrient availability, and internal integrity before committing to DNA synthesis.
S Phase (Synthesis)
The S phase is defined by DNA Synthesis. During this stage, each chromosome is replicated to produce two identical sister chromatids held together at the centromere. DNA replication is semi-conservative and is carried out with high fidelity through coordinated enzymatic complexes. Despite proofreading mechanisms, replication errors can occur, necessitating DNA repair pathways that operate concurrently. Throughout S phase, chromosomes, known as chromatin, remains relatively decondensed to permit access to replication machinery, resulting in a diffuse nuclear appearance under light microscopy.
G2 Phase (Gap 2)
The G2 phase serves as a preparatory interval between DNA replication and mitosis. Although the genome has been duplicated, chromosomes remain in a decondensed state. During G2, the cell synthesizes proteins required for mitotic spindle formation, organelle biogenesis, and cytoskeletal organization. This phase also includes critical checkpoint mechanisms that verify DNA integrity and ensure that replication has been completed accurately prior to entry into mitosis.
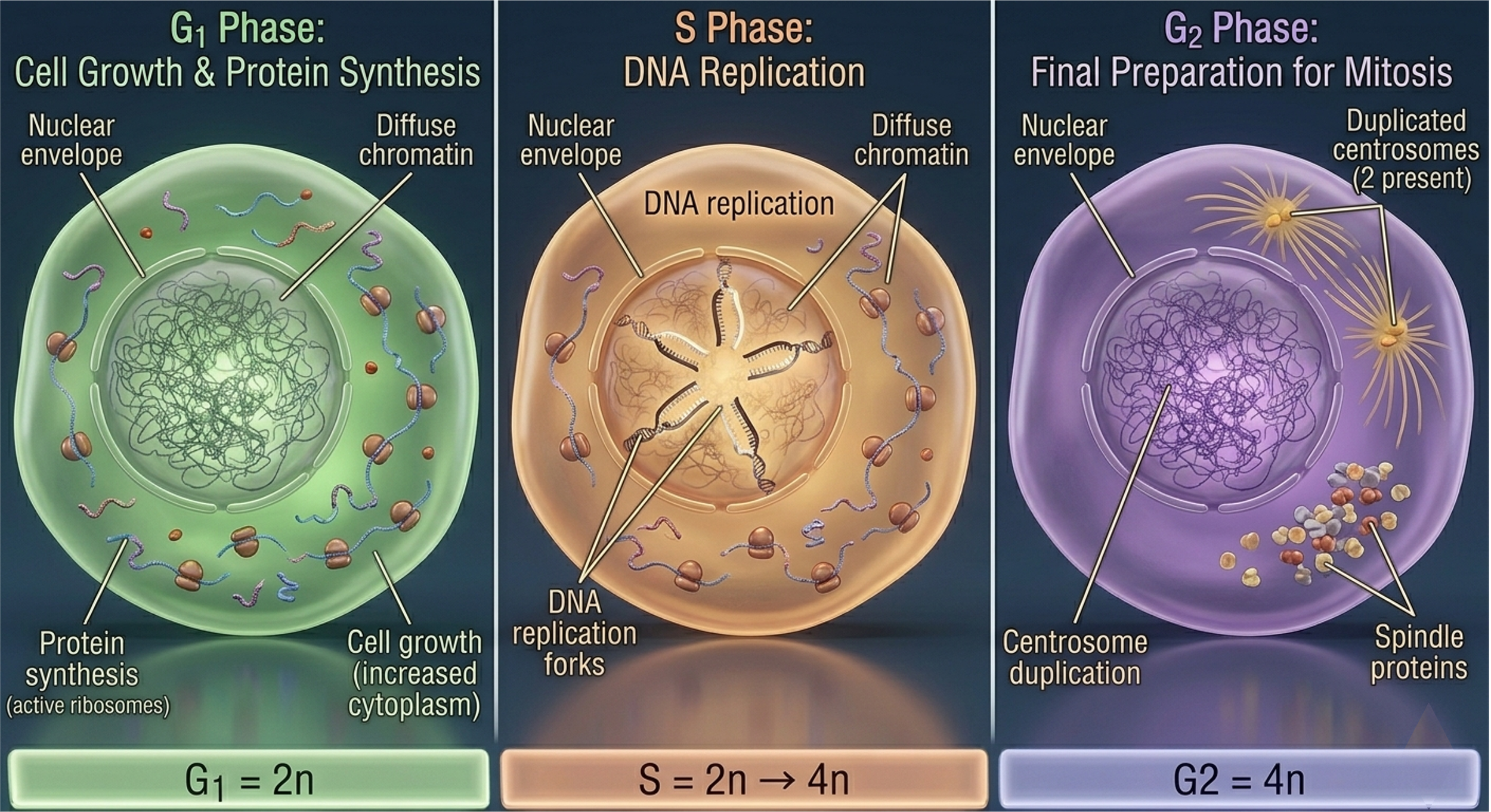
Figure 4. Stages of interphase. G1 involves cellular growth and metabolic activity, S phase includes DNA replication, and G2 prepares the cell for mitosis through protein synthesis and spindle assembly preparation.
Chromosome Architecture During Mitosis
Following interphase, chromatin undergoes progressive condensation into discrete mitotic chromosomes. This compaction facilitates mechanical segregation and reduces entanglement during chromosome movement. Each replicated chromosome consists of two sister chromatids, which are genetically identical copies joined at the centromere. The centromere serves as both a structural cohesion site and a platform for kinetochore assembly. In diploid organisms, chromosomes exist as homologous pairs, with one homolog inherited from each parent. Homologous chromosomes share gene content and structural organization but may differ in allelic composition. It is critical to distinguish that homologous chromosome separation occurs during meiosis I, whereas mitosis involves separation of sister chromatids. The kinetochore is a multiprotein complex assembled at the centromere and serves as the attachment site for spindle microtubules. These microtubules generate mechanical force, enabling chromosome movement and segregation.
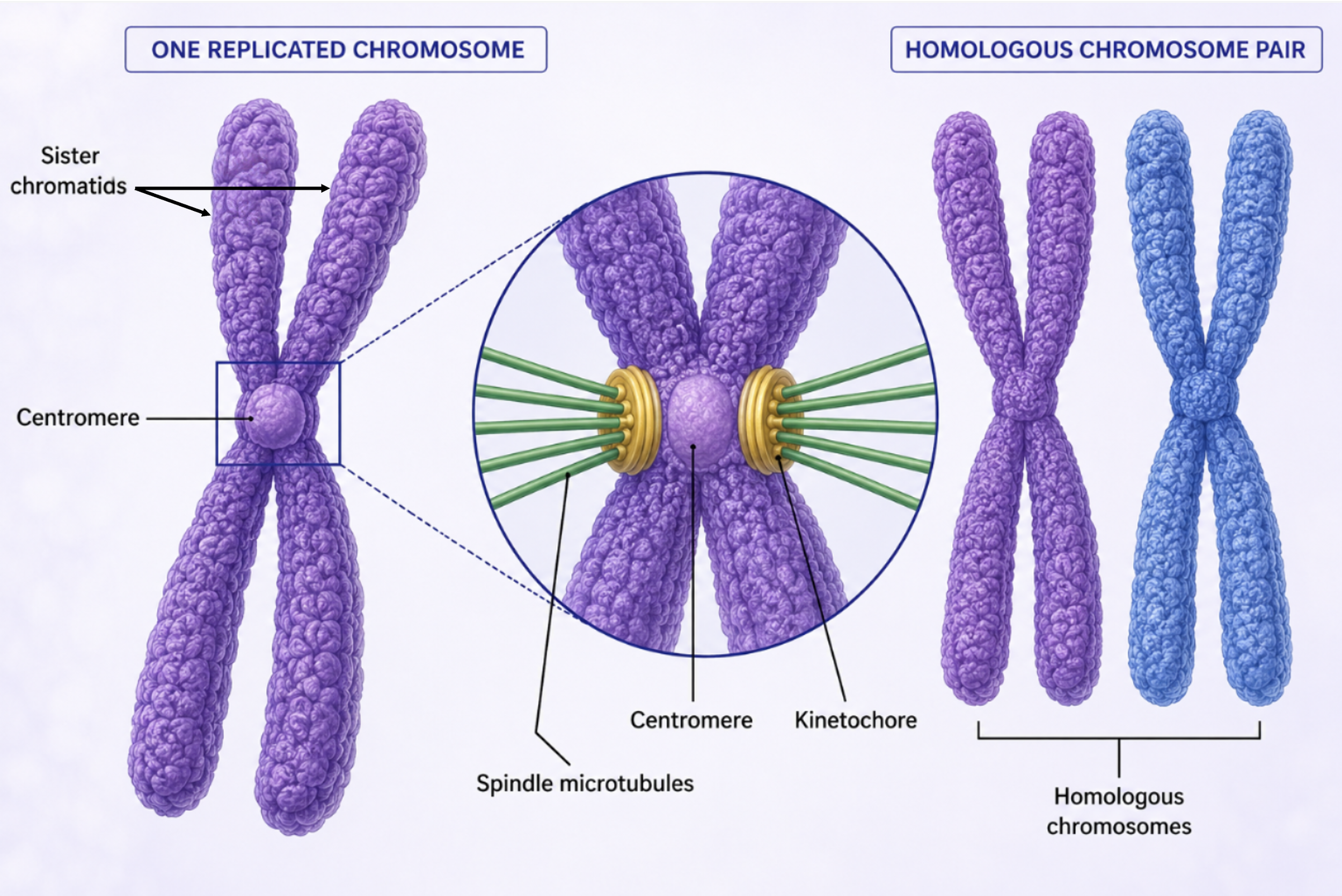
Figure 4. Chromosome Structure. During mitosis, chromatin condenses into chromosomes composed of two sister chromatids joined at the centromere. Kinetochores assembled at the centromere serve as attachment sites for spindle microtubules.
Mitotic Spindle and Cellular Organization
Mitotic chromosome segregation is mediated by the spindle fibers, a dynamic microtubule-based structure organized by centrioles pairs. Centrioles pairs migrate to opposite poles of the cell during prophase and send spindle fibers to the dividing chromosomes. Spindle fibers attach to the kinetochores ensuring that sister chromatids are attached to opposite poles of the cell. This configuration is essential for accurate chromosome segregation and genomic stability.
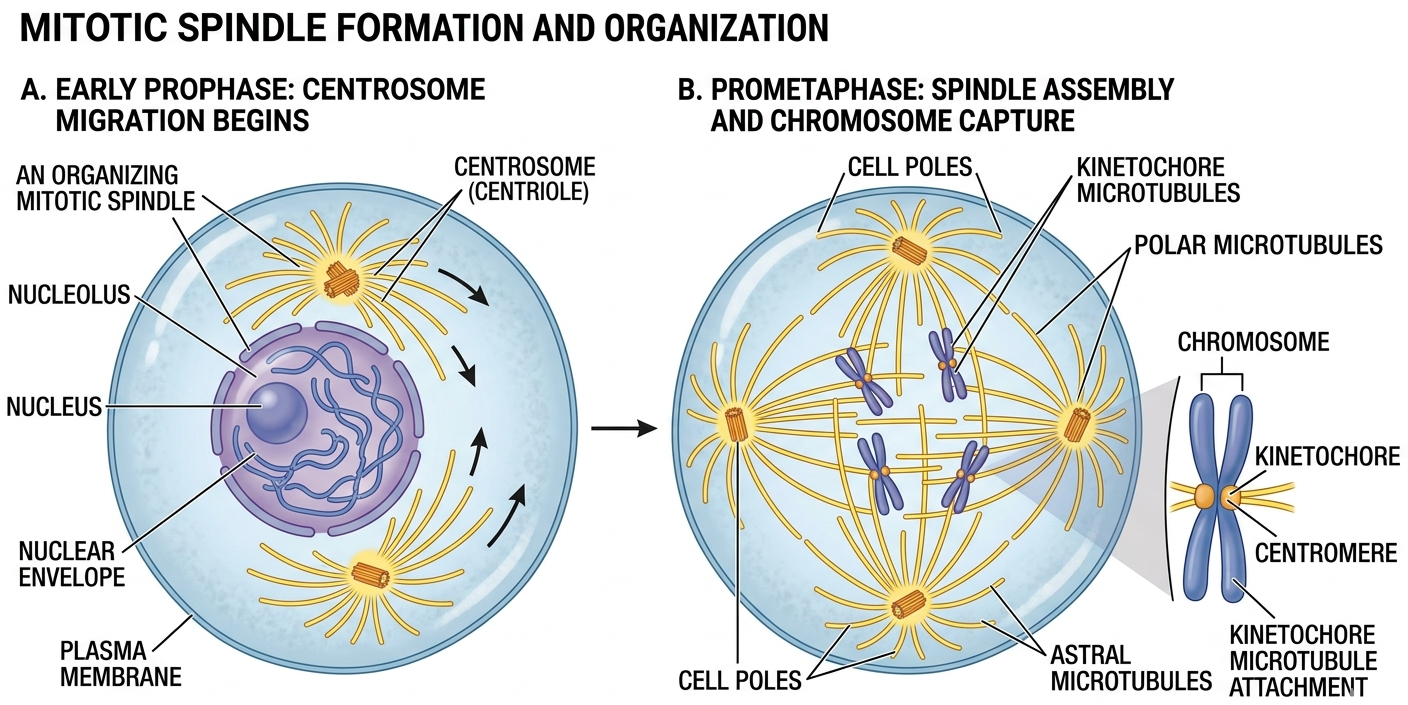
Figure 5. Mitotic Spindles. The mitotic spindle is a dynamic microtubule-based structure that organizes chromosome movement and ensures accurate chromosome segregation during mitosis.
Phases of Mitosis
Mitosis is a continuous process that is conventionally divided into five stages for analytical clarity: prophase, prometaphase, metaphase, anaphase, and telophase. These stages represent progressive structural and functional transitions in chromosome organization and spindle dynamics.
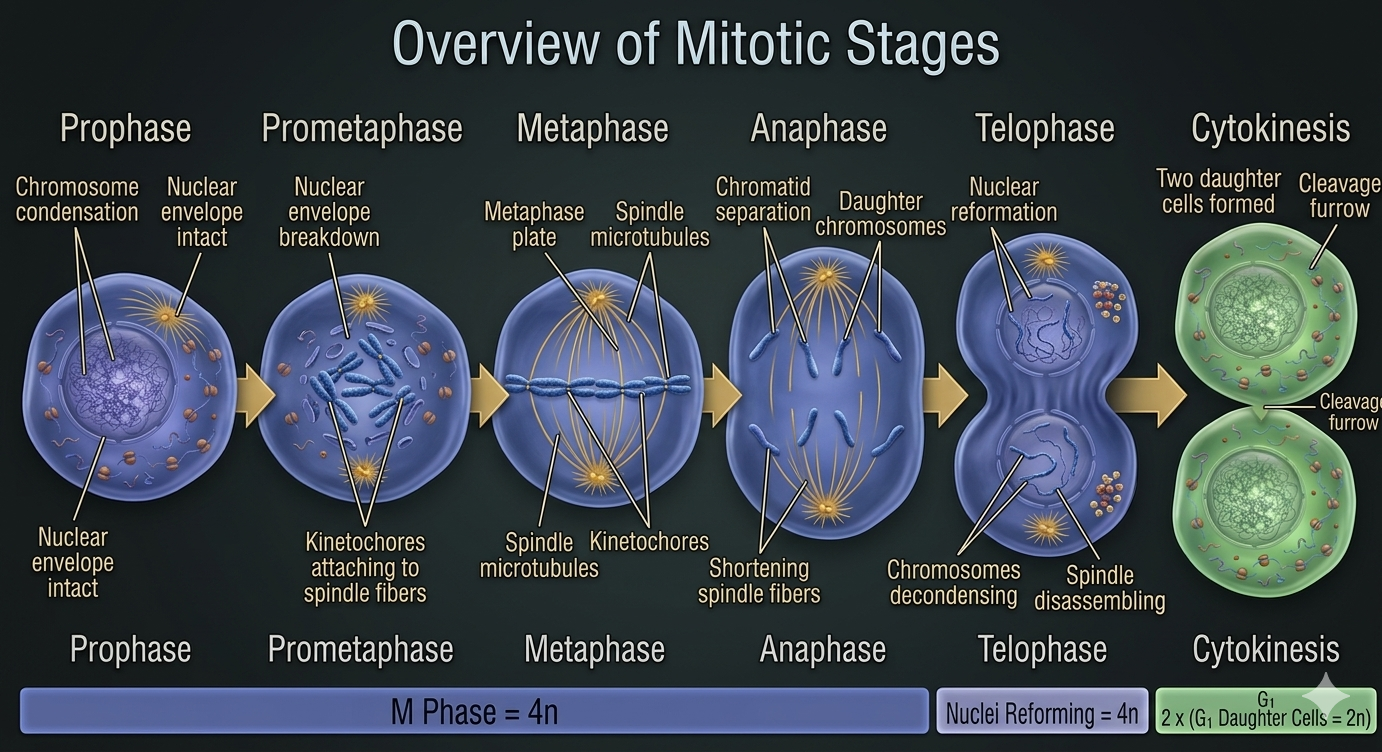
Figure 6. Phase of Mitosis. Mitosis proceeds through a continuous sequence of stages that coordinate chromosome condensation, spindle attachment, chromosome segregation, and nuclear reformation.
Prophase
Mitosis begins with prophase, the first stage in which the cell’s genetic material becomes clearly organized for division. During this stage, chromatin condenses into distinct chromosomes, making them visible under a microscope for the first time in the cell cycle. This condensation is essential because it compacts the DNA into stable structures that can be accurately separated later in mitosis. By the time prophase begins, each chromosome has already been duplicated during the S phase of interphase. As a result, each chromosome consists of two identical copies called sister chromatids, which remain attached at the centromere. In diploid cells, these duplicated chromosomes are arranged as homologous pairs, meaning there are two versions of each chromosome, one inherited from each parent.
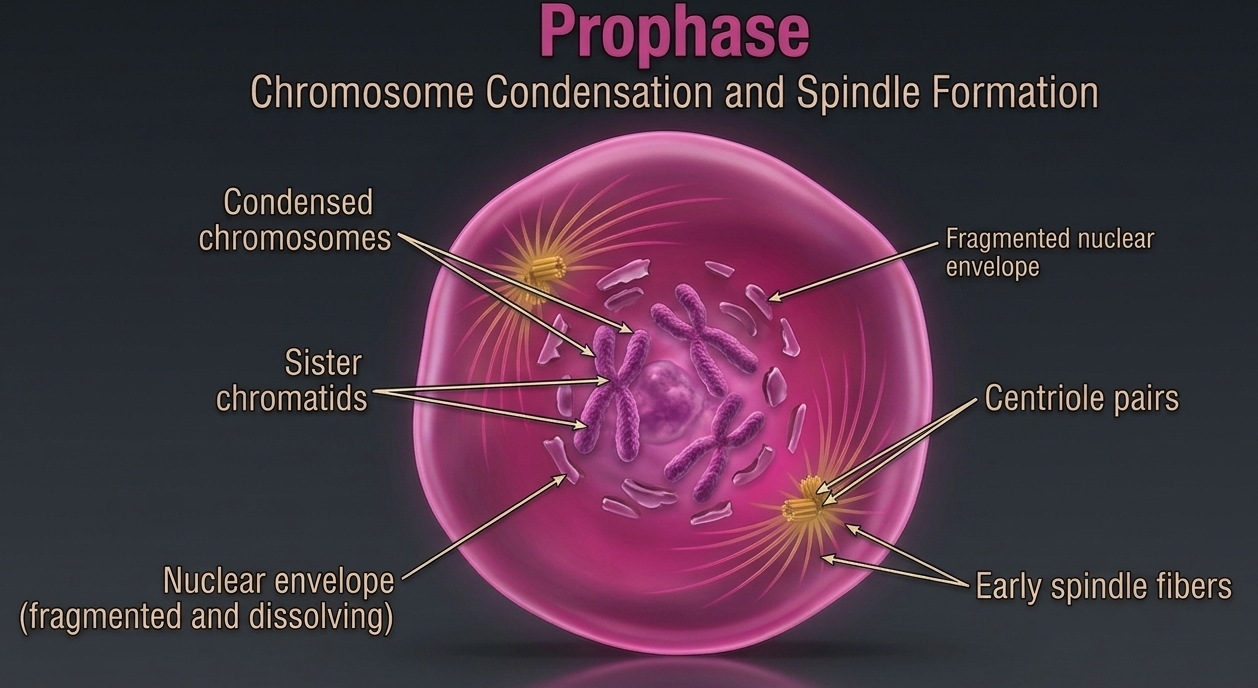
Figure 7. Prophase. During prophase, chromatin condenses into visible duplicated chromosomes composed of two sister chromatids joined at the centromere. This structural organization prepares the genetic material for accurate segregation during mitosis, while homologous chromosome pairs remain present within the diploid cell.
Prometaphase
Prometaphase is often described as a continuation of prophase, but it is also a distinct stage marked by the complete breakdown of nuclear organization and the establishment of direct interactions between chromosomes and the mitotic spindle. During this stage, the nuclear envelope fully disintegrates, removing the physical barrier that previously separated the chromosomes from the rest of the cytoplasm. This breakdown is essential because it allows the mitotic machinery to directly access and manipulate the chromosomes. At the same time, the spindle fibers emerge growing outward from the centriole pairs, which function as organizing centers for the mitotic spindle. As these microtubules elongate, they actively search for and attach to chromosomes. This attachment occurs at specialized protein structures called the kinetochores, which are located on the centromere region of each sister chromatid. Once attachment is established, chromosomes begin to experience mechanical forces that will later align and separate them. Meanwhile, the centriole pairs migrate toward opposite poles of the cell. This movement is critical for ensuring that, when chromosome separation occurs later in mitosis, each daughter cell receives an equal and identical set of genetic material.
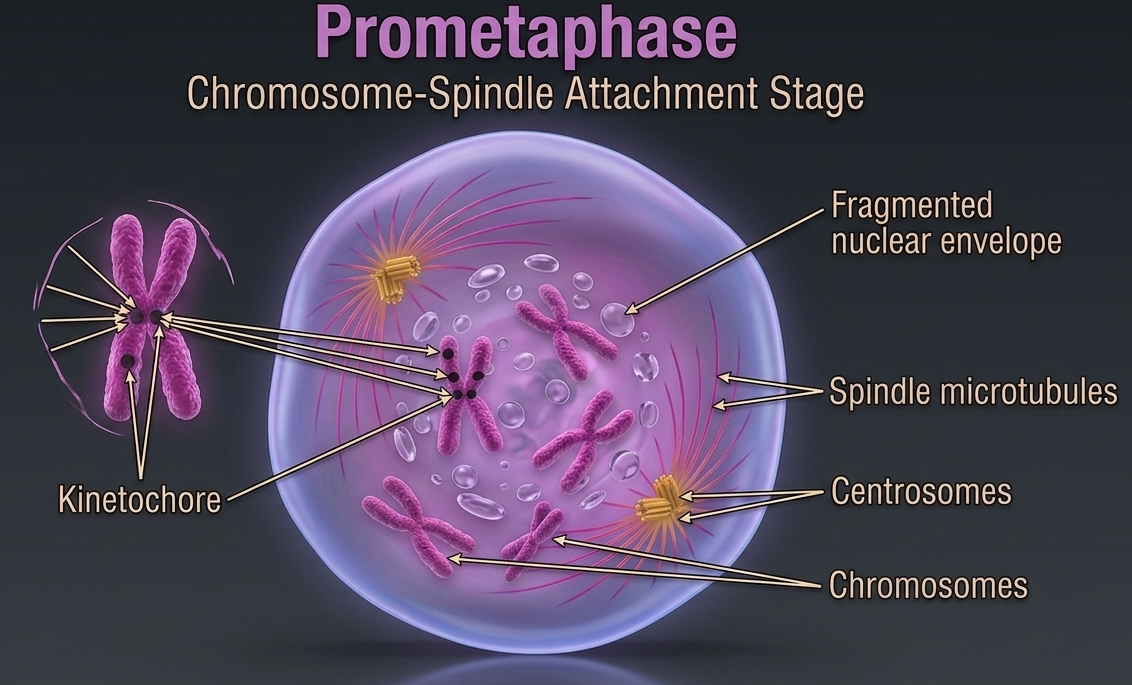
Figure 8. Prometaphase. During prometaphase, the nuclear envelope breaks down and spindle microtubules extend from opposite centrosomes to attach to kinetochores located at the centromeres of sister chromatids. These spindle interactions establish the bipolar mitotic apparatus that will align and separate chromosomes into two genetically identical daughter cells.
Metaphase is the stage of mitosis in which chromosomes reach their most organized and visible alignment. After being captured by spindle fibers during earlier stages, the chromosomes are pulled into a single plane at the center of the cell, known as the metaphase plate. This alignment is not random; it is a controlled checkpoint that ensures each sister chromatid is properly connected before separation occurs. Only when all chromosomes are correctly aligned and under equal tension does the cell proceed to the anaphase, reducing the risk of unequal genetic distribution.
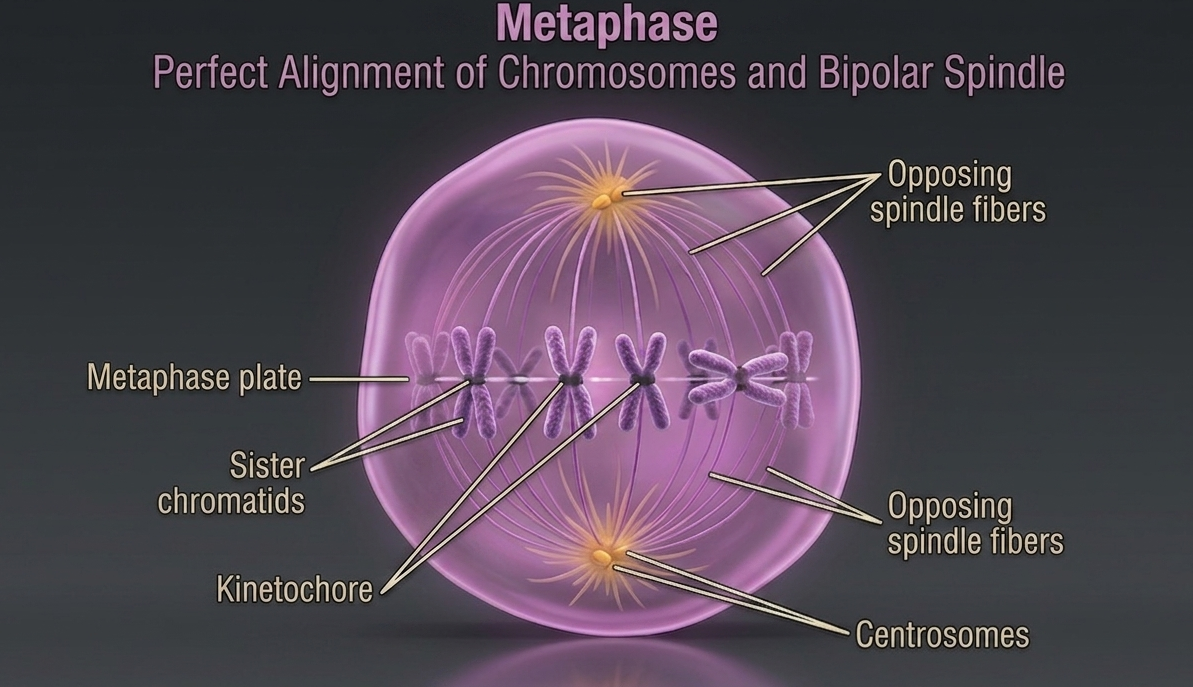
Figure 9. Metaphase. During metaphase, spindle microtubules from opposite poles attach to the kinetochores of sister chromatids and generate tension that aligns the chromosomes along the metaphase plate. This precise arrangement ensures that each daughter cell will receive an identical and complete set of chromosomes during chromosome separation.
Anaphase
Anaphase starts once the chromosomes have aligned at the metaphase plate, the cell is positioned for the transition into chromosome separation. At this point, the spindle apparatus is fully formed, with spindle fibers attached to the kinetochores on each chromosome and extending toward opposite poles of the cell. Spindle microtubules shorten through controlled motor protein activity. As these microtubules shorten, they generate force that pulls the chromosomes away from the metaphase plate toward opposite poles of the cell. This separation process begins when the connections holding the sister chromatids together are broken, allowing each chromatid to move independently. Once separated, each individual sister chromatid is referred to as a daughter chromosome. These daughter chromosomes are genetically identical copies of one another because they were produced through accurate DNA replication during the S phase of interphase. Their equal distribution to opposite sides of the cell ensures that each resulting daughter cell will inherit the same complete set of genetic information.
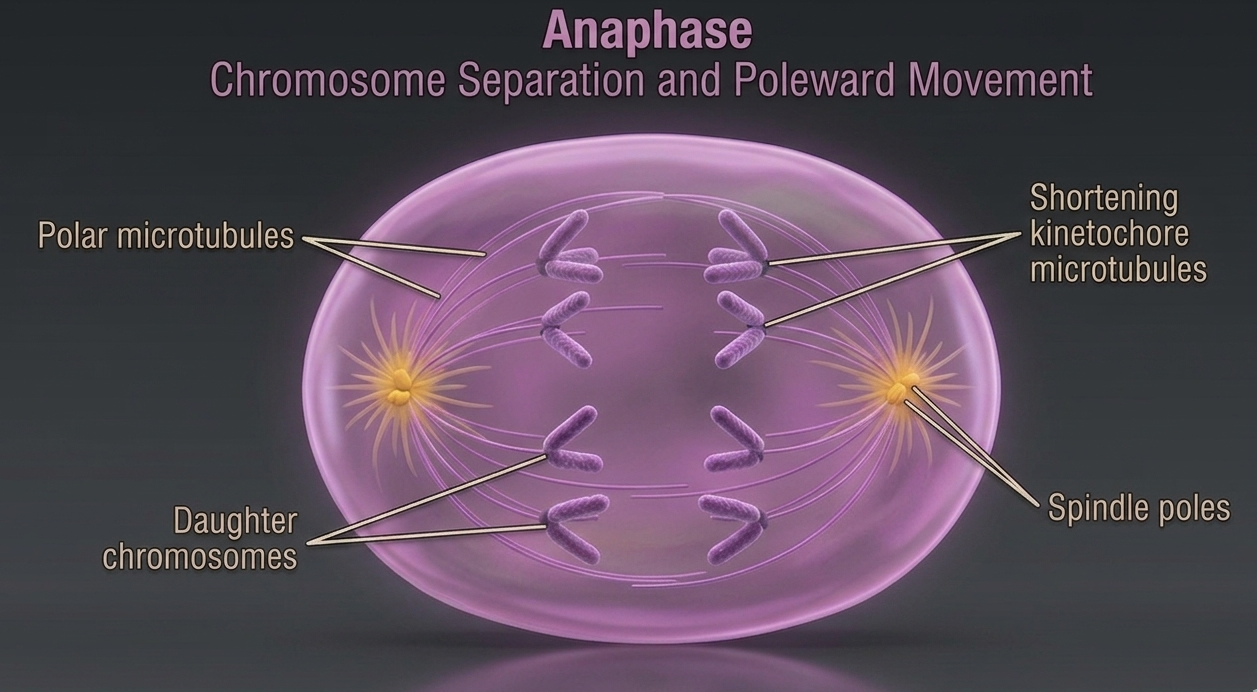
Figure 10. Anaphase. During anaphase, spindle microtubules shorten and pull sister chromatids apart after the centromere connections are released. The separated daughter chromosomes move toward opposite poles of the cell, ensuring equal distribution of identical genetic material to each future daughter cell.
Telophase
During telophase, the cell begins to reverse many of the structural changes that occurred during earlier stages of mitosis. The spindle fibers disassemble as they are no longer needed for chromosome movement, and the mitotic spindle gradually disappears. At this point, the daughter chromosomes have already reached opposite poles of the cell and are fully separated into two distinct groups. The visible chromosomes relax, once again becoming chromatin. As the chromosomes arrive at their respective poles, the new nuclear envelopes reform around each set of chromosomes, reconstructing two separate nuclear compartments within the same cell. After the nuclear envelope reforms during telophase, the chromosomes begin to decondense back into chromatin, returning to a less compact state that allows normal cellular activity and gene expression to resume. By the end of telophase, the cell contains two nuclei within a single cell body, referred to as daughter nuclei. Each nucleus contains an identical and complete set of chromosomes, ensuring that the genetic information has been accurately and equally distributed in preparation for the final stage of cell division, cytokinesis.
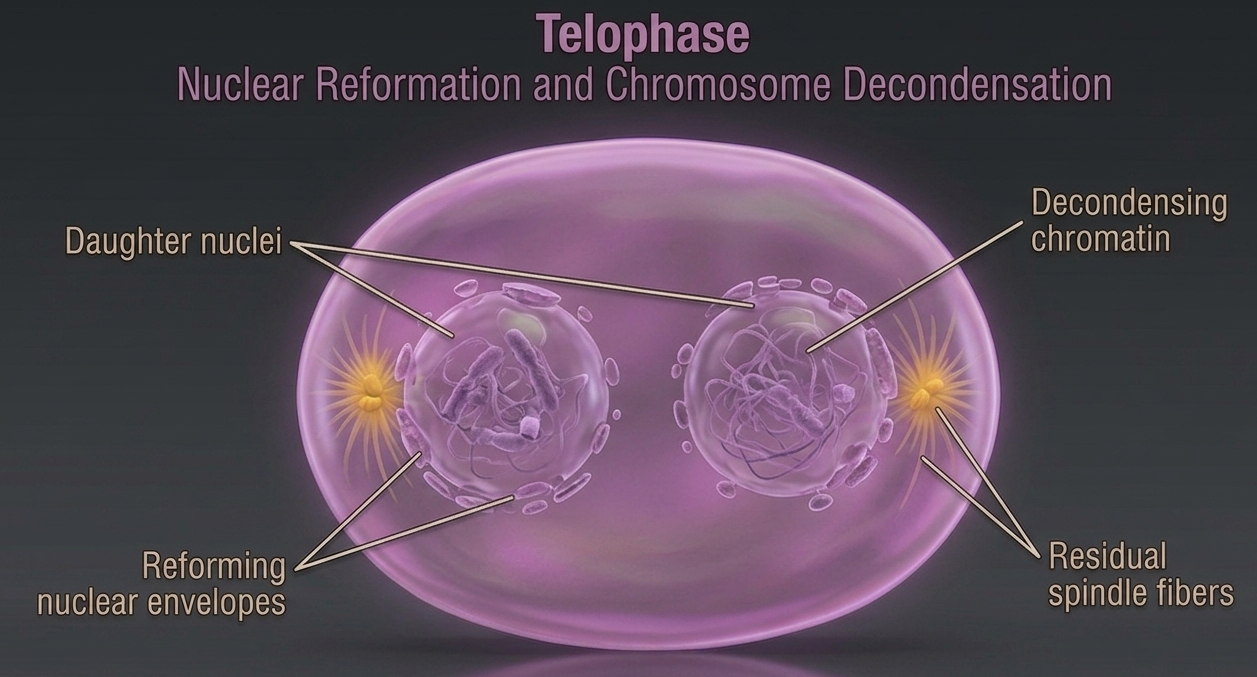
Figure 11. Telophase. During telophase, daughter chromosomes reach opposite poles of the cell, spindle fibers disassemble, and new nuclear envelopes reform around each chromosome set. The chromosomes then decondense back into chromatin, producing two genetically identical daughter nuclei in preparation for cytokinesis.
Cytokinesis: Cytoplasmic Division
At this point, the cell completes division through a process known as cytokinesis, in which the cytoplasm divides to produce two separate daughter cells. In animal cells, cytokinesis occurs through the formation of a cleavage furrow. This furrow develops due to a contractile ring made primarily of actin filaments (along with myosin motor proteins) positioned just beneath the plasma membrane. As this ring contracts, it progressively tightens, pulling the membrane inward like a drawstring and eventually pinching the cell into two separate daughter cells. In plant cells, cytokinesis occurs differently because of the rigid cell wall. Instead of forming a cleavage furrow, vesicles from the Golgi apparatus accumulate at the center of the cell along the equatorial plane. These vesicles fuse to form a structure called the cell plate, which gradually expands outward. The cell plate contains materials that will become the new cell wall. As it grows, it eventually fuses with the existing plasma membrane on both sides, completing the physical separation of the two daughter cells.
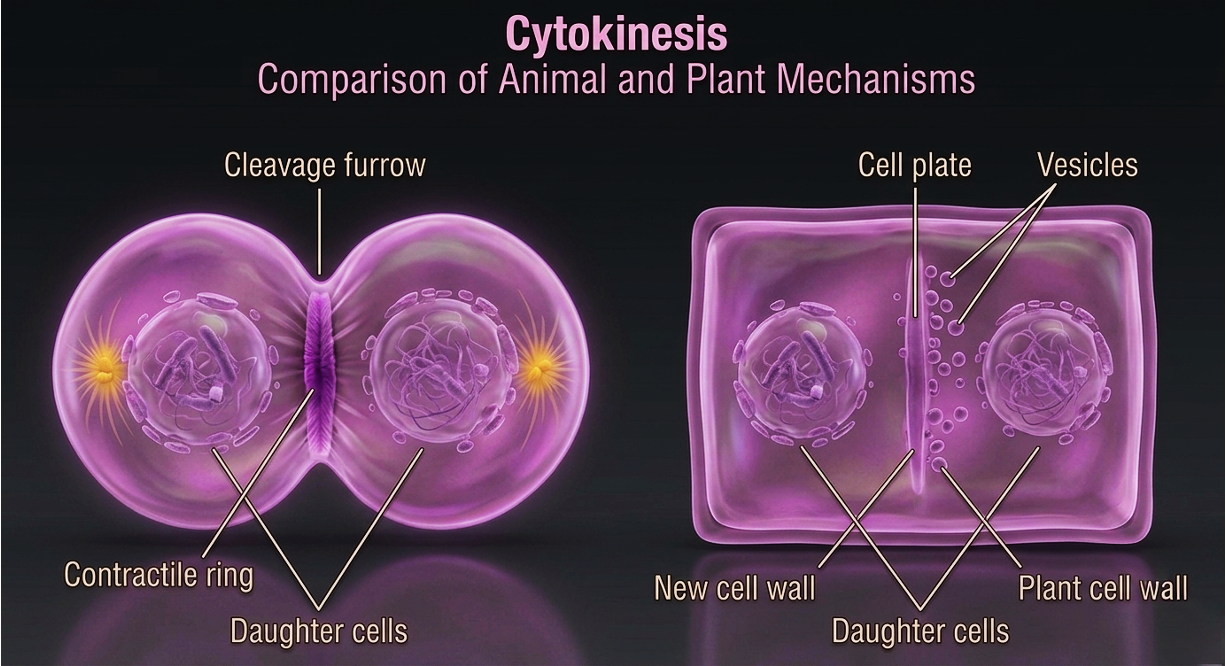
Figure 12. Cytokinesis. During cytokinesis, the cytoplasm divides to form two separate daughter cells. Animal cells complete this process through contraction of an actin-based cleavage furrow, whereas plant cells form a cell plate from Golgi-derived vesicles that develops into a new cell wall between the daughter cells.
Biological Significance of Cell Division
Cell Division in eukaryotes enable reproduction, growth, tissue maintenance, and regeneration. The cell cycle begins when a single fertilized egg, and drives the development to a complex multicellular organism. The cell cycle ensure that tissues remain functional over time by replacing cells lost to damage, wear, or programmed cell death. These processes also support repair and regeneration. The extent of this regenerative capacity varies across tissues depending on stem cell availability and regulatory control mechanisms. A key function of mitosis and cytokinesis is the preservation of genomic continuity across somatic cell lineages. Accurate DNA replication during interphase and precise chromosome segregation during mitosis ensure that each daughter cell receives an identical set of genetic information. This maintains stable gene expression and cellular function across generations of cells. At a broader systems level, cell division underlies developmental biology, tissue homeostasis, and the long-term stability of multicellular organisms. Proper regulation maintains a balance between cell proliferation and cell loss. Disruption of this balance can result in degenerative disease or uncontrolled growth such as cancer.
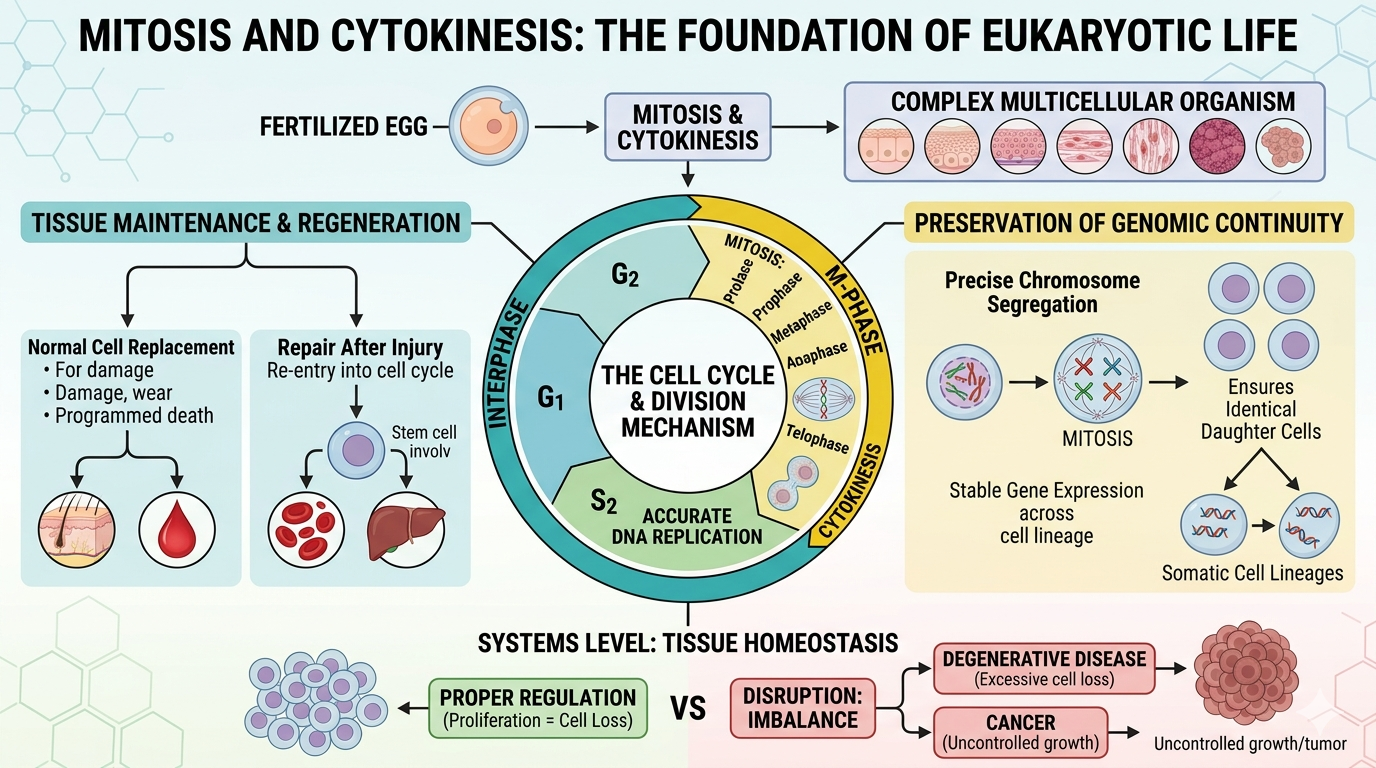
Figure 13. The Cell Cycle. Mitosis and cytokinesis are essential processes of eukaryotic life that enable growth, tissue maintenance, repair, and the development of multicellular organisms. By accurately replicating and dividing genetic material, these processes preserve genomic continuity and maintain tissue stability, while failures in cell cycle regulation can contribute to diseases such as cancer.