
Chapter: The Cell
The Cell Theory
The cell theory is one of the foundational theories of biology. A scientific theory is a well-tested and strongly supported explanation of natural phenomena that is supported by extensive evidence collected through the scientific method. Modern biology rests heavily on two major theories: the theory of evolution by natural selection and the cell theory. The cell theory explains the basic nature of living organisms and states that: (1) all living organisms are composed of cells, (2) the cell is the basic unit of life, and (3) all cells arise from preexisting cells. Together, these principles unify all fields of biology because every known living organism is cellular in structure and depends on cellular processes to survive.
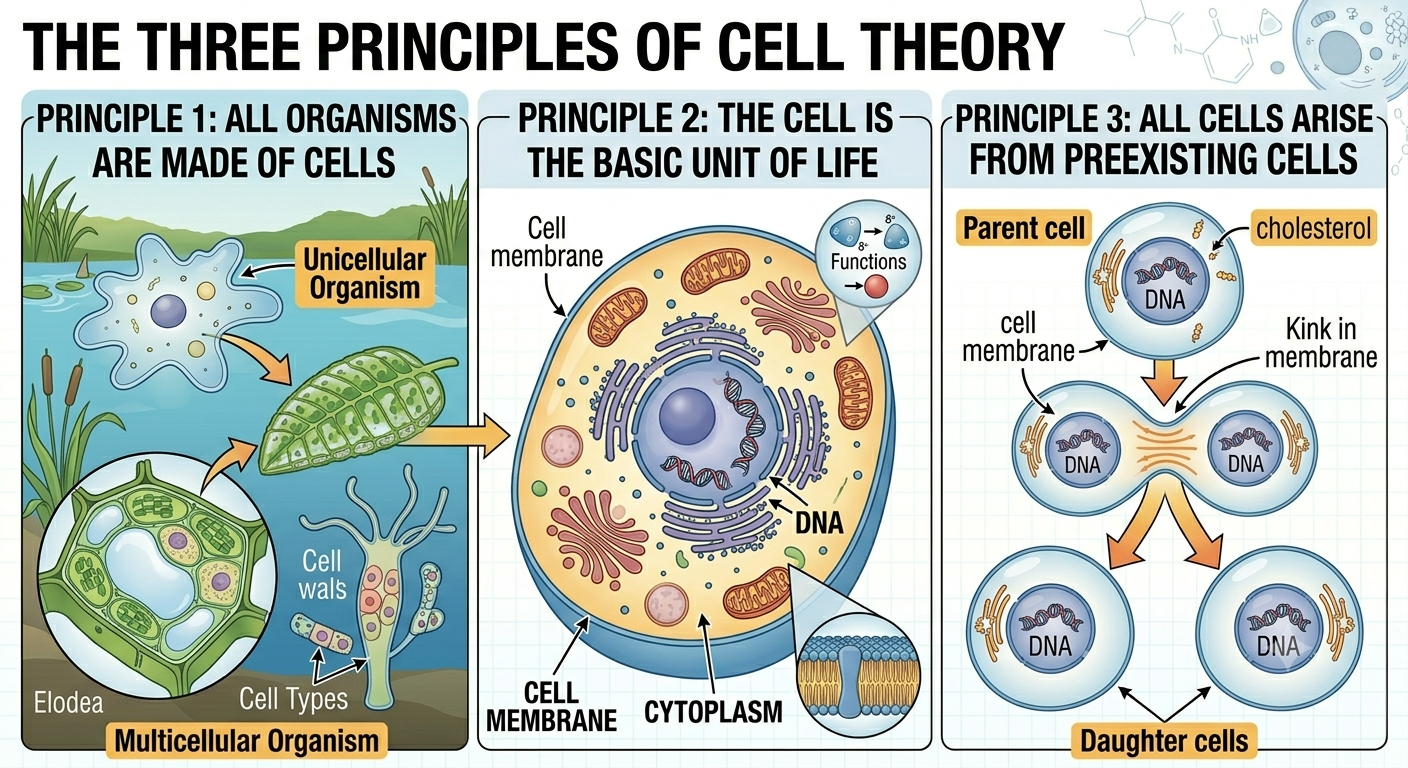
Figure 1. The three principles of cell theory. Cell theory states that all living organisms are composed of cells, the cell is the basic unit of life, and all cells arise from preexisting cells through cell division. These principles unify all biology because every known living organism depends on cellular structure and cellular processes.
The Cell is the Basic Unit of Life
A cell is the smallest structure capable of carrying out all functions necessary for life. Every cell is enclosed by a cell membrane, also known as the plasma membrane, which separates the internal environment of the cell from the external environment. This membrane maintains homeostasis, meaning the cell can regulate internal conditions such as pH, ion concentration, water balance, and nutrient levels despite changes in the outside environment. Although many biochemical reactions can be reproduced in a laboratory outside of cells, living cells organize and coordinate these reactions with remarkable precision and efficiency. Cells obtain and use energy, synthesize proteins, store genetic information, respond to stimuli, and reproduce. Structures smaller than cells, such as proteins, DNA, or viruses, may participate in life processes, but they are not considered independently alive because they cannot perform all characteristics of life on their own.
All Living Organisms Are Composed of Cells
All known living organisms are made of one or more cells. Some organisms, such as bacteria and many protists, are unicellular, meaning a single cell carries out all life functions. Other organisms, including plants, animals, and fungi, are multicellular, consisting of many specialized cells working together as tissues and organs. The cell theory emphasizes that no alternative unit of life exists outside the cell. This principle has generated debate regarding viruses, which possess genetic material and can evolve, reproduce, and respond to environmental conditions, yet lack a true cell membrane and cannot reproduce independently without infecting a host cell. Because viruses depend entirely on living cells for replication and metabolism, most biologists do not classify them as fully living organisms.
All Cells Come From Preexisting Cells
The third principle of the cell theory states that all cells arise from other cells through the process of cell division. This concept, known as biogenesis, replaced the earlier belief in spontaneous generation, which proposed that living organisms could emerge directly from nonliving matter. For centuries, many people believed that maggots spontaneously formed from decaying meat or that fleas emerged from dust. In the nineteenth century, the French scientist Louis Pasteur performed experiments demonstrating that microorganisms originated from existing microorganisms rather than appearing spontaneously. His work provided strong evidence that life comes only from preexisting life and helped establish modern microbiology.
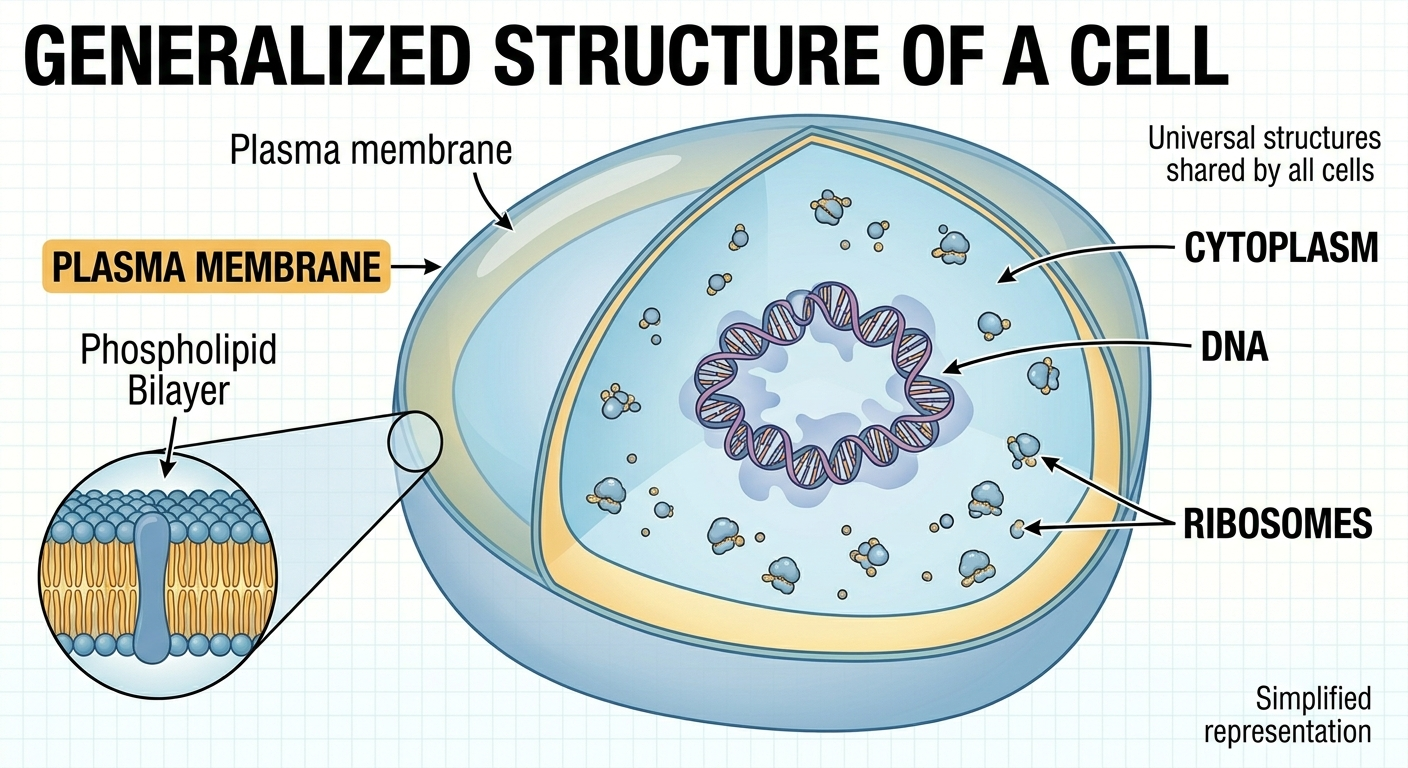
Figure 2. Structures shared by all cells. All cells possess a plasma membrane, cytoplasm, ribosomes, and DNA. These universal features support metabolism, protein synthesis, homeostasis, and inheritance in every known form of life.
Characteristics Shared by All Cells
Although cells vary greatly in size and complexity, all cells share several fundamental characteristics. Every cell is surrounded by a cell membrane composed primarily of a phospholipid bilayer and proteins. This membrane creates a controlled internal environment filled with a gel-like fluid called the cytoplasm. All cells also contain DNA, the hereditary molecule that stores genetic information. DNA carries instructions for producing RNA, and RNA directs the synthesis of proteins at structures called ribosomes. This flow of genetic information is summarized by the central dogma of molecular biology, which states that information generally moves from DNA → RNA → protein. Proteins then perform most structural and biochemical functions within the cell. One remarkable discovery in biology is that nearly all organisms use the same genetic code. A gene from one organism can often function inside another organism because the codons of DNA and RNA specify the same amino acids in nearly all life forms. This universal code provides strong evidence that all life on Earth descended from a common ancestor.
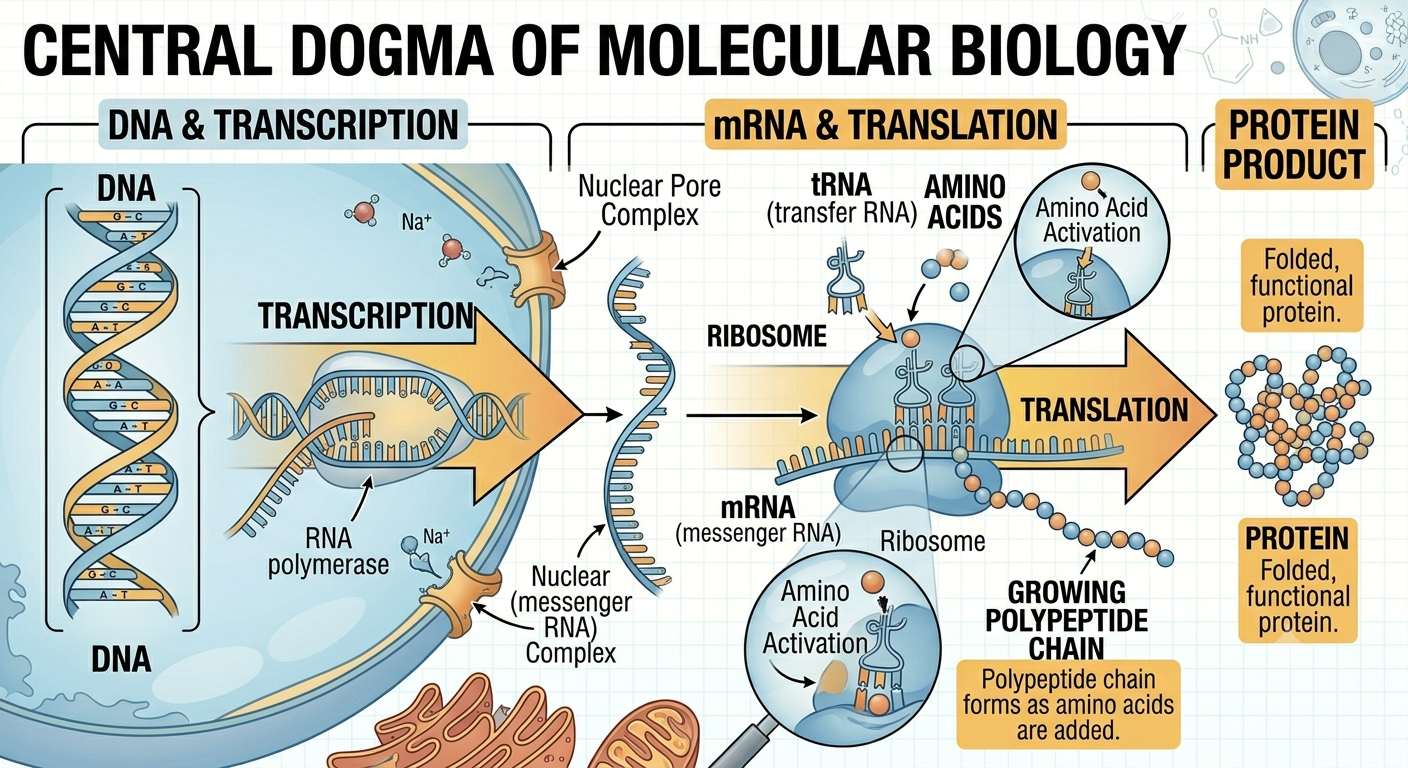
Figure 3. The central dogma of molecular biology. Genetic information is generally transferred from DNA to RNA through transcription and from RNA to protein through translation. Proteins then carry out most structural and biochemical functions within cells.
The Three Domains of Life
Modern classification divides all cellular life into three major groups called domains: Bacteria, Archaea, and Eukarya. These domains represent the broadest categories of biological classification and are based largely on genetic evidence. Organisms in the domains Bacteria and Archaea are classified as prokaryotes, meaning they lack a membrane-bound nucleus and membrane-bound organelles. Organisms in the domain Eukarya are known as eukaryotes and possess a true nucleus and numerous organelles enclosed by membranes. Although bacteria and archaea appear superficially similar under a microscope, molecular evidence shows that archaea are genetically more closely related to eukaryotes than to bacteria. One major difference is that bacterial cell walls contain the carbohydrate peptidoglycan, whereas archaea and eukaryotes lack peptidoglycan in their cell structures.
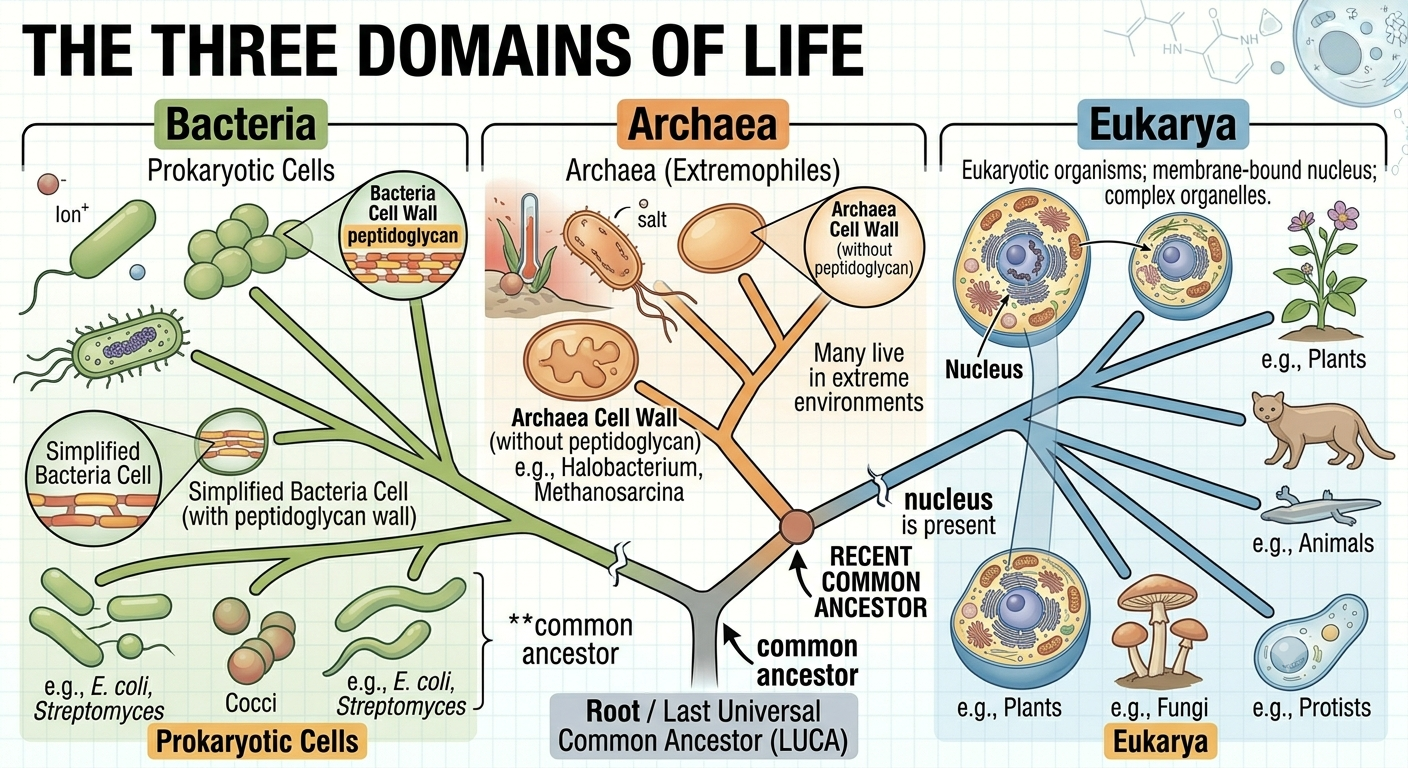
Figure 4. The three domains of life. Modern organisms are classified into the domains Bacteria, Archaea, and Eukarya based largely on genetic evidence. Molecular data indicate that archaea are more closely related to eukaryotes than to bacteria despite their superficial structural similarities.
Prokaryotic Cells
Prokaryotic cells are generally smaller and simpler than eukaryotic cells. They lack a true nucleus, so their DNA exists in a concentrated region called the nucleoid rather than inside a membrane-bound compartment. Prokaryotic DNA is usually organized as a single circular chromosome. Although prokaryotes lack membrane-bound organelles, they still contain ribosomes, cytoplasm, and a plasma membrane. Many bacteria also possess a rigid cell wall that provides structural support and protection. Most prokaryotes are unicellular, though some species form colonies or exhibit simple multicellular behaviors. In prokaryotes, transcription and translation occur simultaneously because there is no nucleus separating DNA from ribosomes. As RNA is being synthesized from DNA, ribosomes can immediately begin translating the RNA into protein.
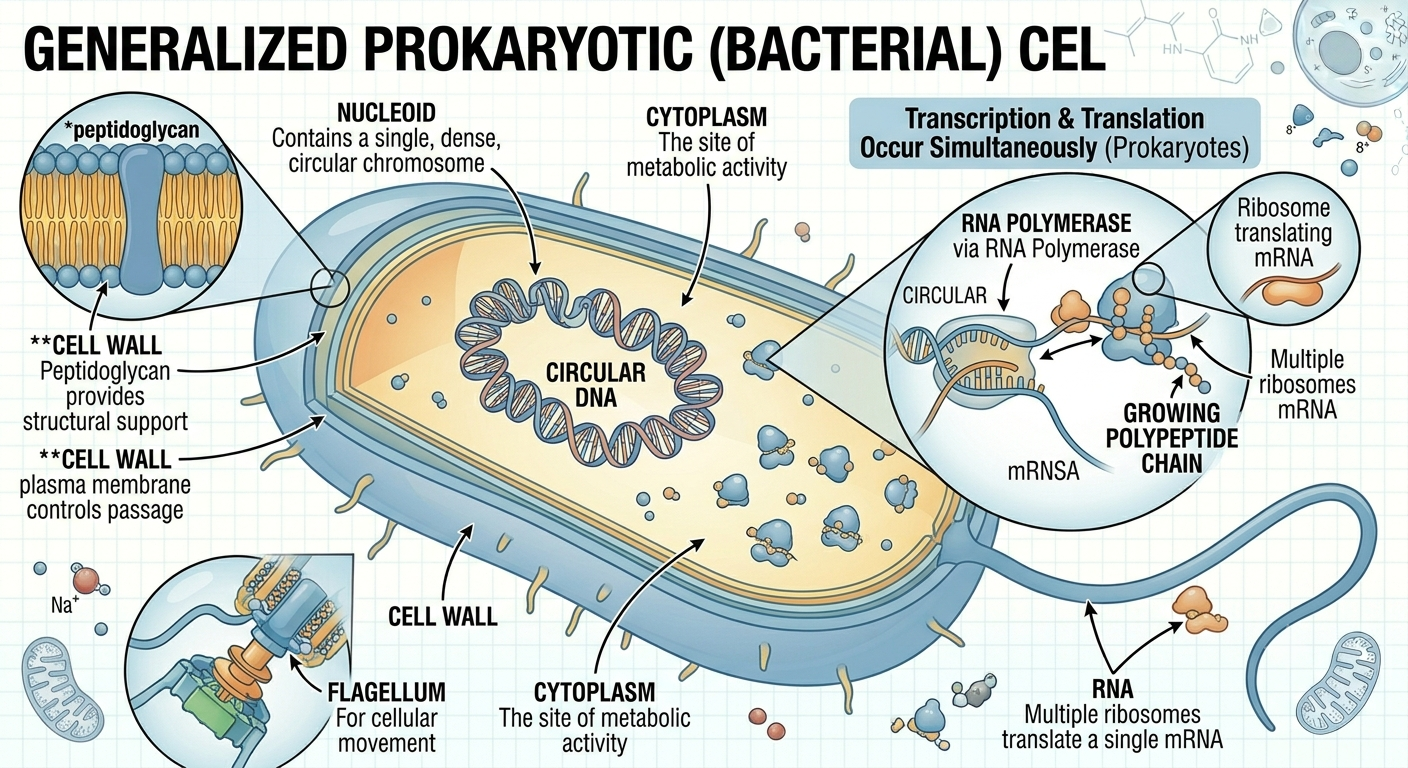
Figure 5. Structure of a prokaryotic cell. Prokaryotes lack a membrane-bound nucleus and membrane-bound organelles. Their DNA is located within a nucleoid region, and transcription and translation occur simultaneously within the cytoplasm.
Eukaryotic Cells
Eukaryotic cells are larger and more structurally complex than prokaryotic cells. Their DNA is enclosed within a membrane-bound nucleus, and they contain numerous specialized organelles that compartmentalize cellular functions. Eukaryotic DNA is organized into multiple linear chromosomes rather than a single circular chromosome. Eukaryotes include both unicellular organisms, such as many protists, and multicellular organisms, including plants, animals, and fungi. Compartmentalization allows eukaryotic cells to carry out many biochemical processes simultaneously and with greater efficiency. The evolution of membrane-bound organelles was a major step in the history of life because it allowed increased cellular specialization and complexity.
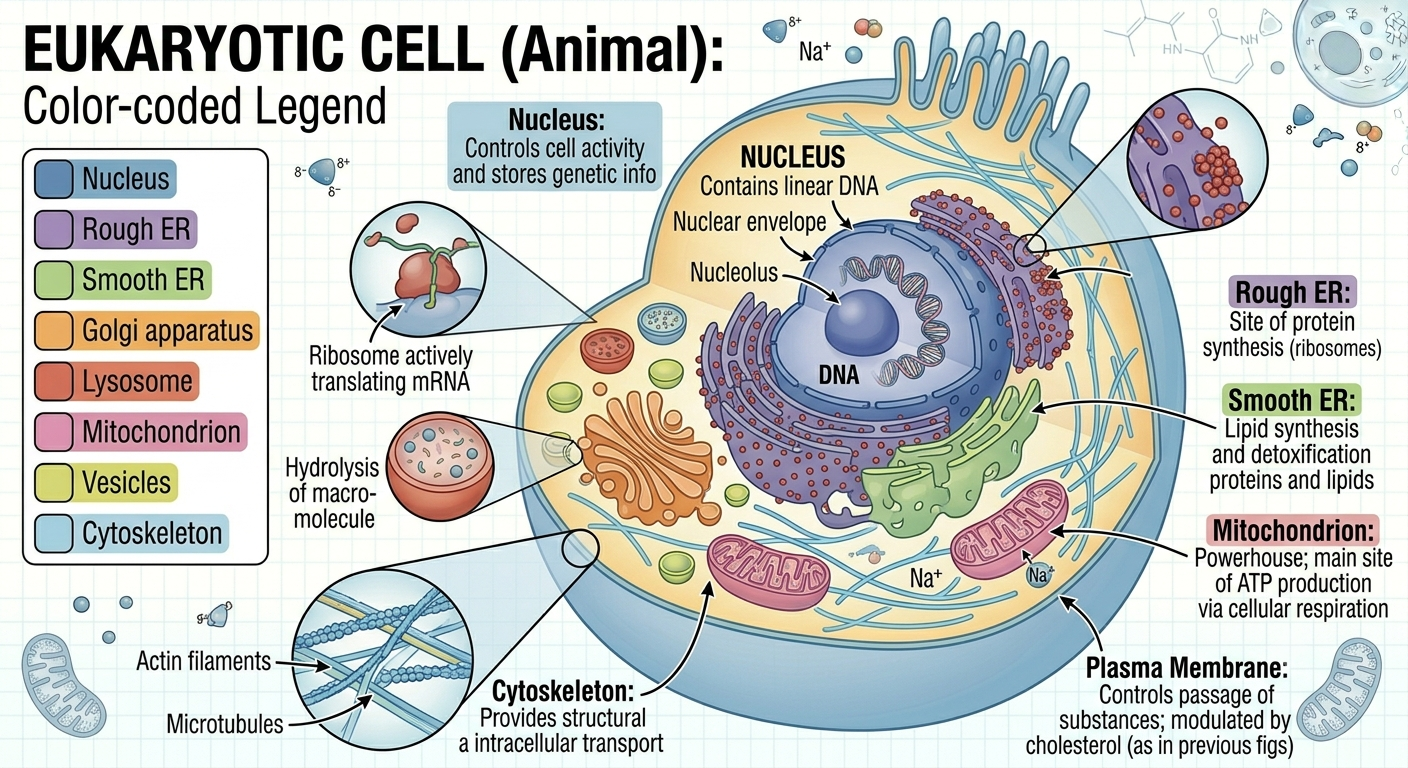
Figure 6. Structure of a eukaryotic cell. Eukaryotic cells contain membrane-bound organelles that compartmentalize cellular processes. This internal organization increases efficiency and allows greater cellular specialization and complexity.
Tour through a Eukaryotic Cell
Organelles Involved in Protein Synthesis
Eukaryotic cells make proteins using a step-by-step process involving several organelles. DNA in the nucleus stores the instructions and is used to make mRNA through transcription. The mRNA is then edited and sent out of the nucleus to ribosomes on the rough endoplasmic reticulum or in the cytosol. At the ribosome proteins are synthesized through translation. After a protein is made, it is migrates to the ends of the rough endoplasmic reticulum, and packaged into transport vesicles. These vesicles are transported on the cytoskeleton network to the cis-face of the Golgi apparatus for where the protein is modified, stored and eventually shipped out of the trans-face of the Golgi apparatus through another transport vesicle. From there, proteins are delivered to their final destinations, such as the cell membrane or outside the cell. Damaged proteins are later broken down and recycled by lysosomes.
Other organelles
Some organelles in eukaryotic cells are not involved in protein synthesis but serve other essential functions. Mitochondria produce ATP through cellular respiration, making energy from glucose; eukaryotic cells can produce up to about 38 ATP per glucose, while prokaryotes produce about 2 ATP because they lack mitochondria. The smooth endoplasmic reticulum (smooth ER) makes lipids such as membrane fats and steroid hormones and also helps detoxification and calcium storage. Vacuoles function as storage sacs, storing water and other materials; they are large in plant cells to maintain structure (turgor pressure) and smaller in animal cells for temporary storage of nutrients, waste, and water.
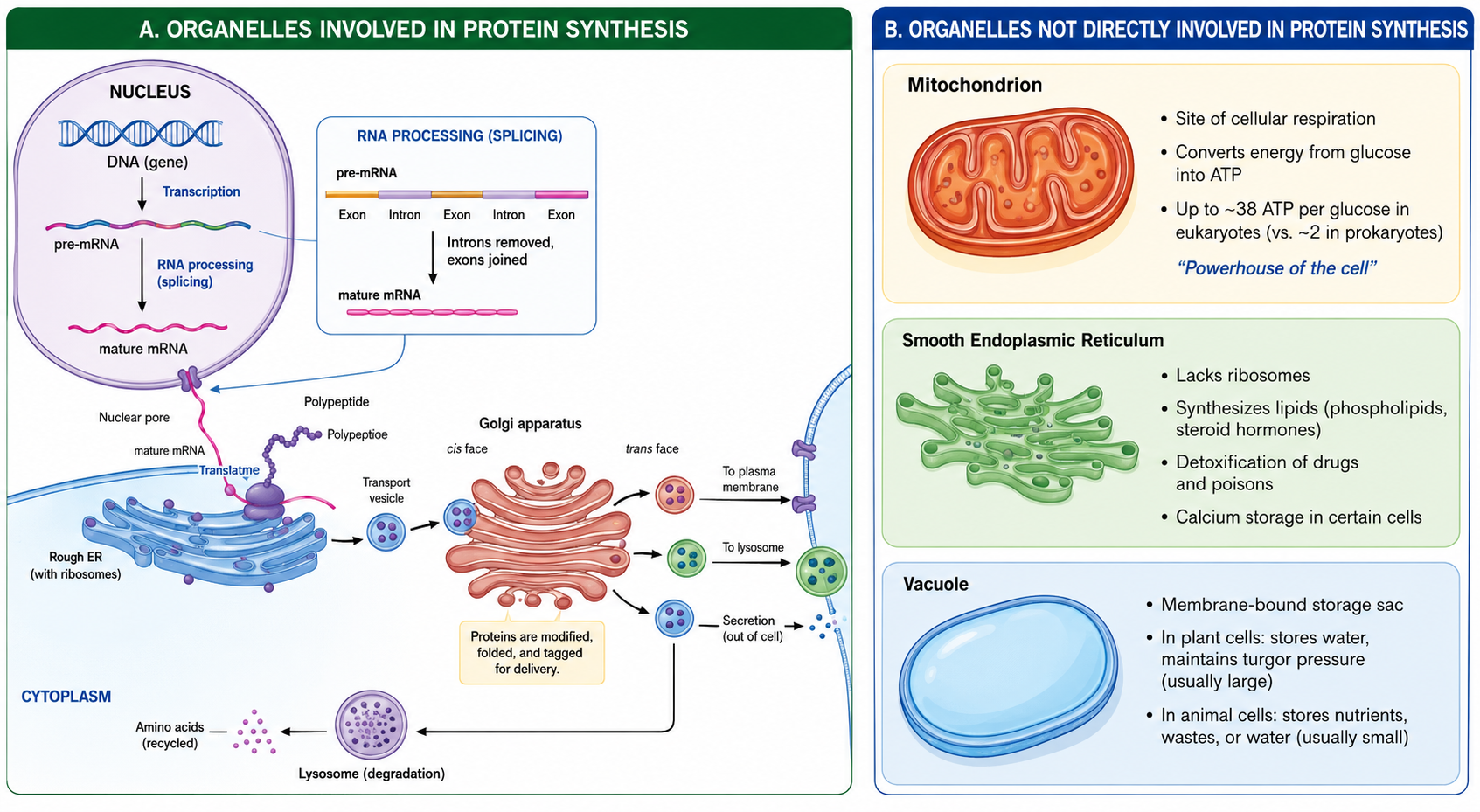
Figure 7. The complete lifecycle of a protein in a eukaryotic cell. Genetic information stored in DNA is transcribed and translated into a protein, which is processed through the endomembrane system. The coordinated activity of the nucleus, rough endoplasmic reticulum, Golgi apparatus, vesicles, and lysosomes ensures that proteins are correctly synthesized, modified, and delivered to their functional destinations.
The Origin of the Endomembrane System
One important feature of eukaryotic cells is the endomembrane system, which includes the nucleus, endoplasmic reticulum, Golgi apparatus, vesicles, and lysosomes. Scientists believe this system evolved through inward foldings of the ancestral cell membrane. Over time, these membrane infoldings became separated from the plasma membrane and formed internal compartments. This process likely produced the nuclear envelope surrounding the DNA and eventually gave rise to the interconnected membrane systems within modern eukaryotic cells. The development of internal membranes greatly increased cellular efficiency by separating incompatible biochemical reactions into specialized compartments.
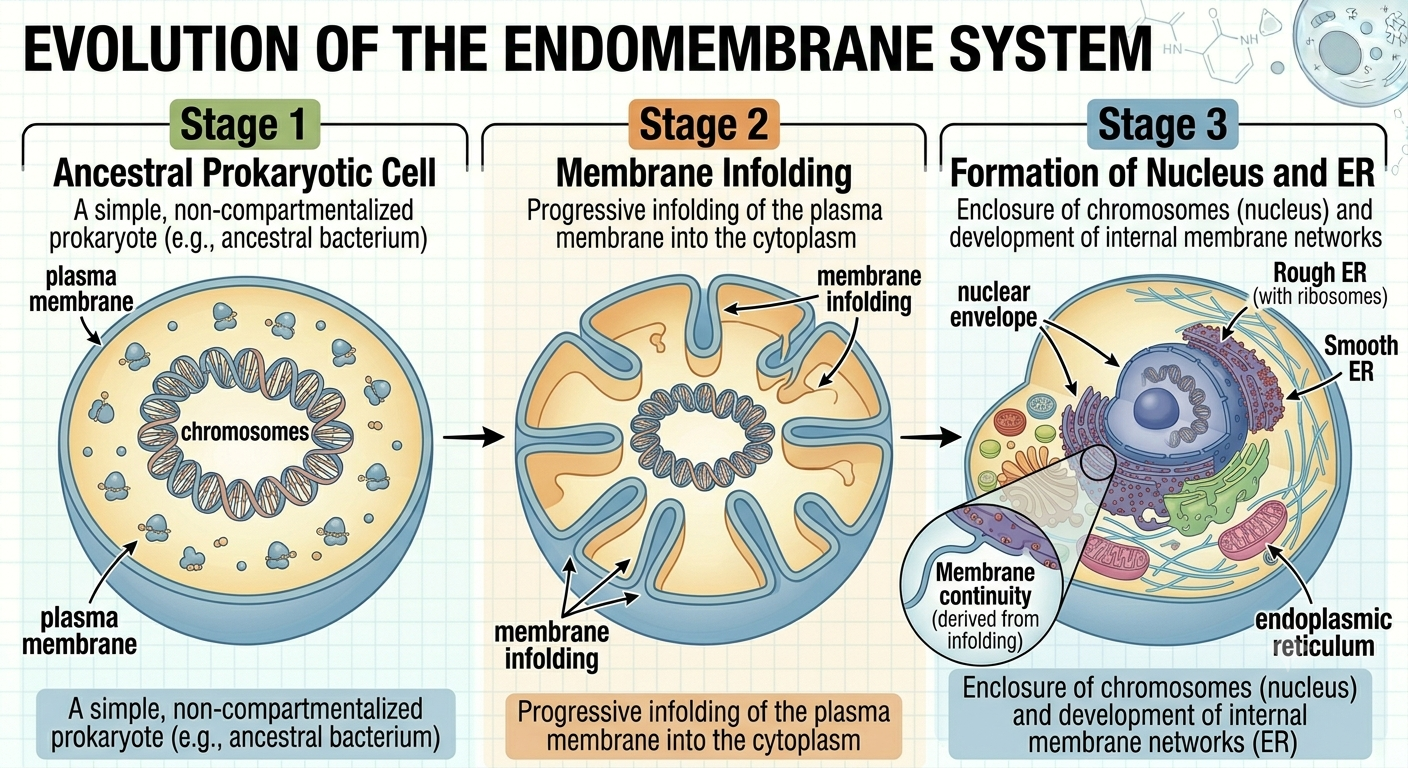
Figure 8. Proposed origin of the endomembrane system. Internal membrane systems likely evolved through infoldings of the ancestral plasma membrane. These infoldings eventually formed the nucleus and interconnected organelles of the endomembrane system.
The Nucleus
The nucleus is the defining organelle of eukaryotic cells. It is enclosed by a double membrane called the nuclear envelope, which contains openings known as nuclear pores that regulate movement of molecules into and out of the nucleus. Inside the nucleus, DNA is organized into structures called chromosomes. The nucleus controls cellular activities because it contains the genetic instructions for protein synthesis. Within the nucleus is a dense region called the nucleolus, where ribosomal RNA and ribosomal subunits are produced. One major advantage of the nucleus is that it separates transcription from translation, allowing eukaryotic cells to regulate gene expression more precisely through RNA processing before proteins are synthesized.
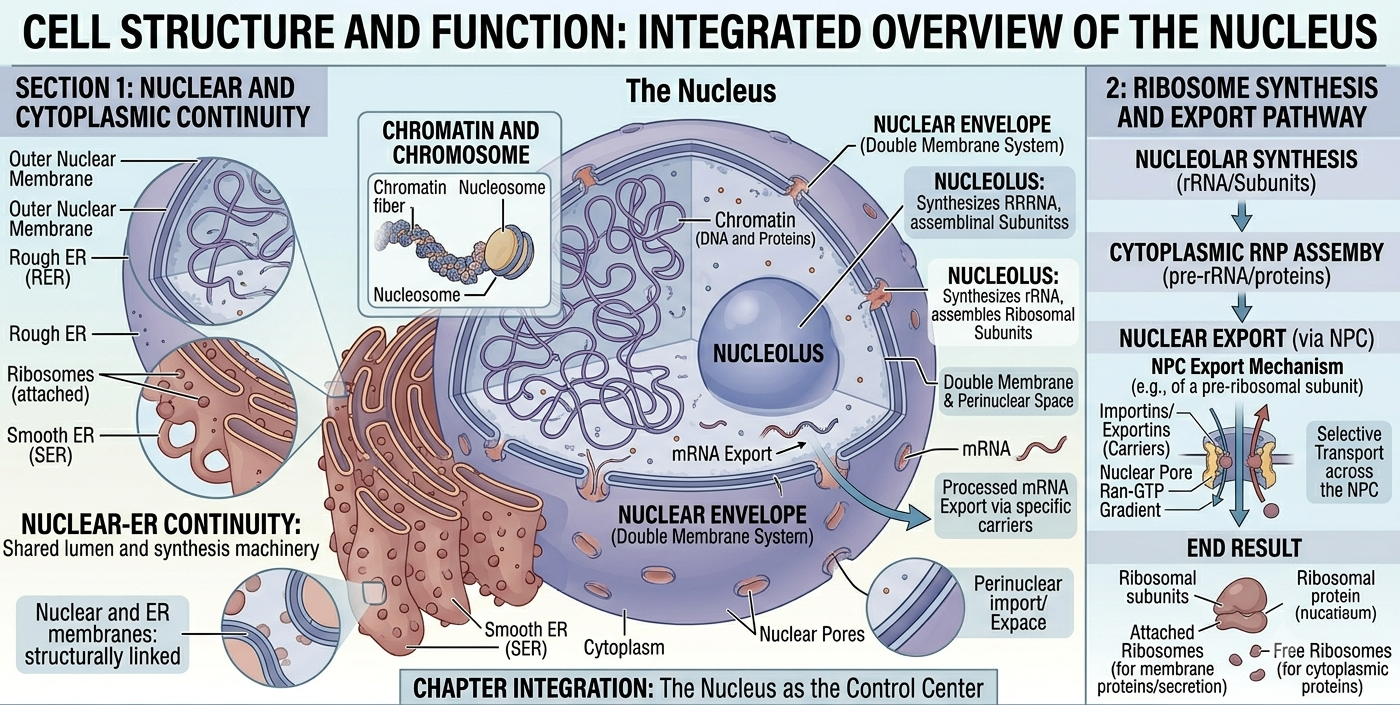
Figure 9. Structure of the nucleus. The nucleus houses the genetic material of eukaryotic cells and separates transcription from translation. Nuclear pores regulate molecular transport between the nucleus and cytoplasm.
The Endoplasmic Reticulum
The endoplasmic reticulum (ER) is an extensive network of interconnected membranes continuous with the nuclear envelope. The rough endoplasmic reticulum is covered with ribosomes and functions primarily in protein synthesis and protein modification. Proteins synthesized on the rough ER are often destined for secretion, insertion into membranes, or use within lysosomes. The smooth endoplasmic reticulum lacks ribosomes and functions mainly in lipid synthesis, detoxification, and calcium storage. Together, the rough and smooth ER play central roles in the production and processing of biological molecules.
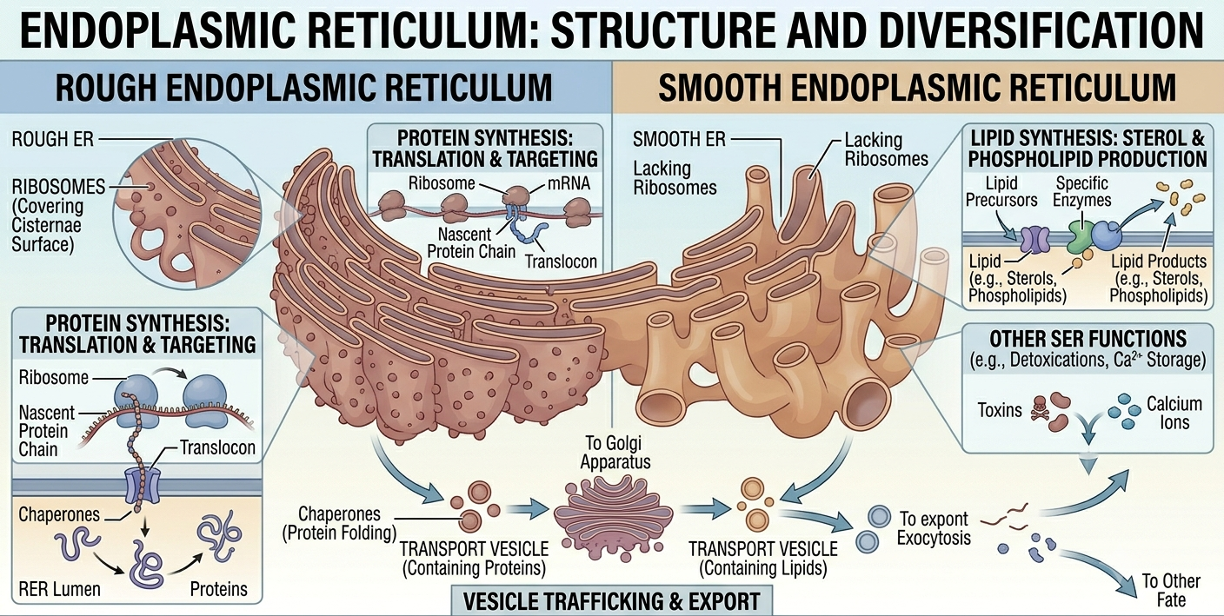
Figure 10. Rough and smooth endoplasmic reticulum. The rough ER synthesizes and processes proteins using attached ribosomes, whereas the smooth ER functions primarily in lipid synthesis, detoxification, and calcium storage.
The Golgi Apparatus
The Golgi apparatus modifies, sorts, and packages proteins and lipids produced by the endoplasmic reticulum. It consists of flattened membrane sacs called cisternae. Transport vesicles carrying newly synthesized proteins fuse with the receiving side of the Golgi, where proteins are chemically modified, folded, and tagged for delivery. Processed proteins are then packaged into vesicles and transported to their final destinations, including the plasma membrane, lysosomes, or outside the cell through exocytosis. The Golgi apparatus functions much like a cellular shipping and distribution center.Vesicles and Cellular Transport
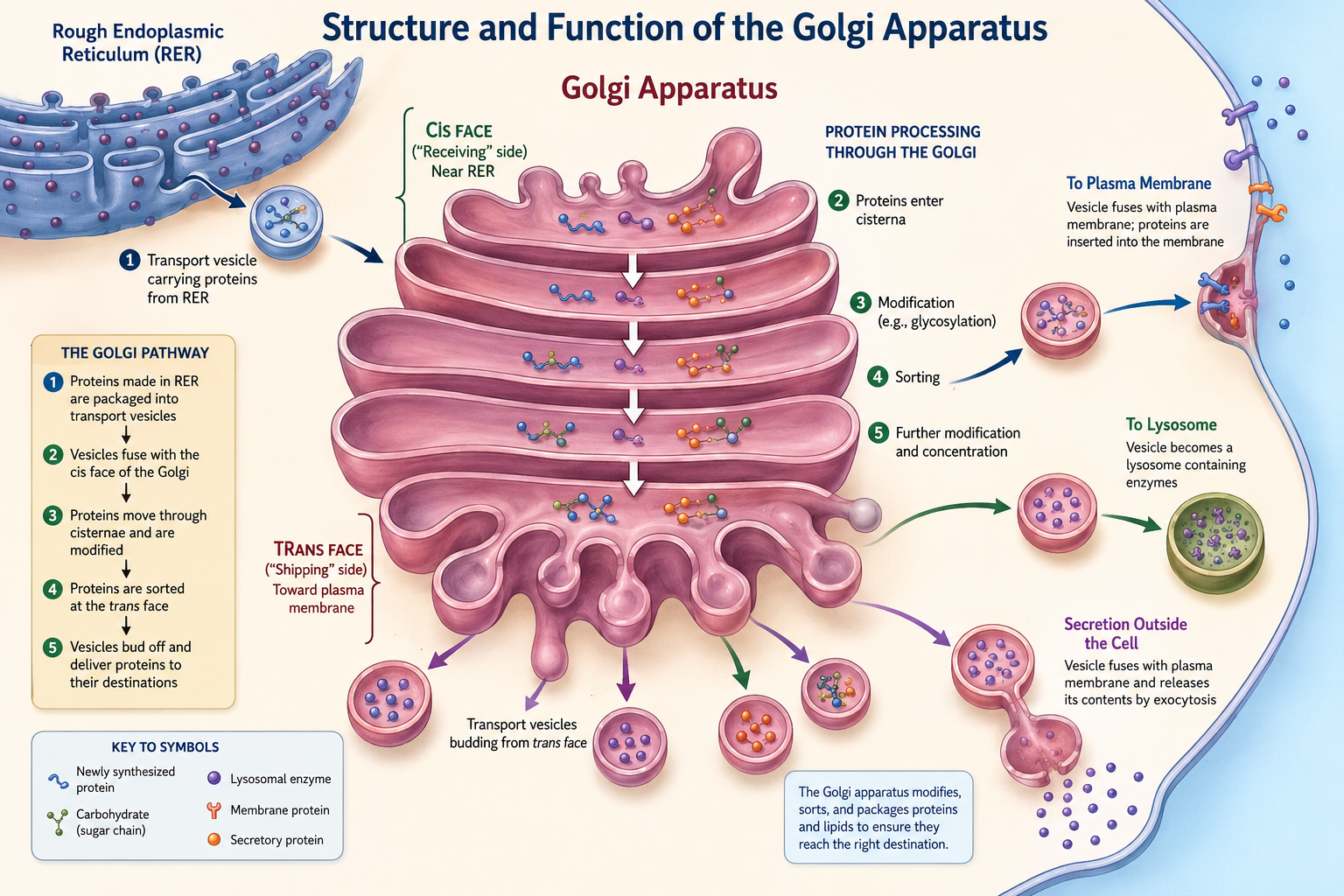
Figure 11. Structure and function of the Golgi apparatus. Newly synthesized proteins produced by the rough endoplasmic reticulum are transported to the Golgi apparatus in vesicles. Within the Golgi, proteins are chemically modified, sorted, and packaged into new vesicles that deliver them to lysosomes, the plasma membrane, or outside the cell through exocytosis. The Golgi apparatus functions as the cellular processing and shipping center.
Vesicles
Vesicles are small membrane-bound sacs that transport substances within cells and between cells and their environment. Vesicles form from portions of cellular membranes that pinch inward or outward. In endocytosis, the plasma membrane folds inward and engulfs materials from outside the cell, forming a vesicle inside the cytoplasm. In exocytosis, vesicles fuse with the plasma membrane and release their contents outside the cell. Vesicles also transport proteins between organelles such as the rough ER and Golgi apparatus. Because vesicles are enclosed by phospholipid bilayers, they can merge with other membranes efficiently and selectively.
Vacuoles
Vacuoles are specialized vesicles primarily involved in storage. Plant cells typically possess a very large central vacuole that stores water, ions, pigments, nutrients, and waste products. The central vacuole also helps maintain turgor pressure, which supports the structure of plant tissues. Animal cells may contain smaller vacuoles that perform storage and transport functions. By isolating substances from the cytoplasm, vacuoles help regulate cellular chemistry and maintain internal balance.
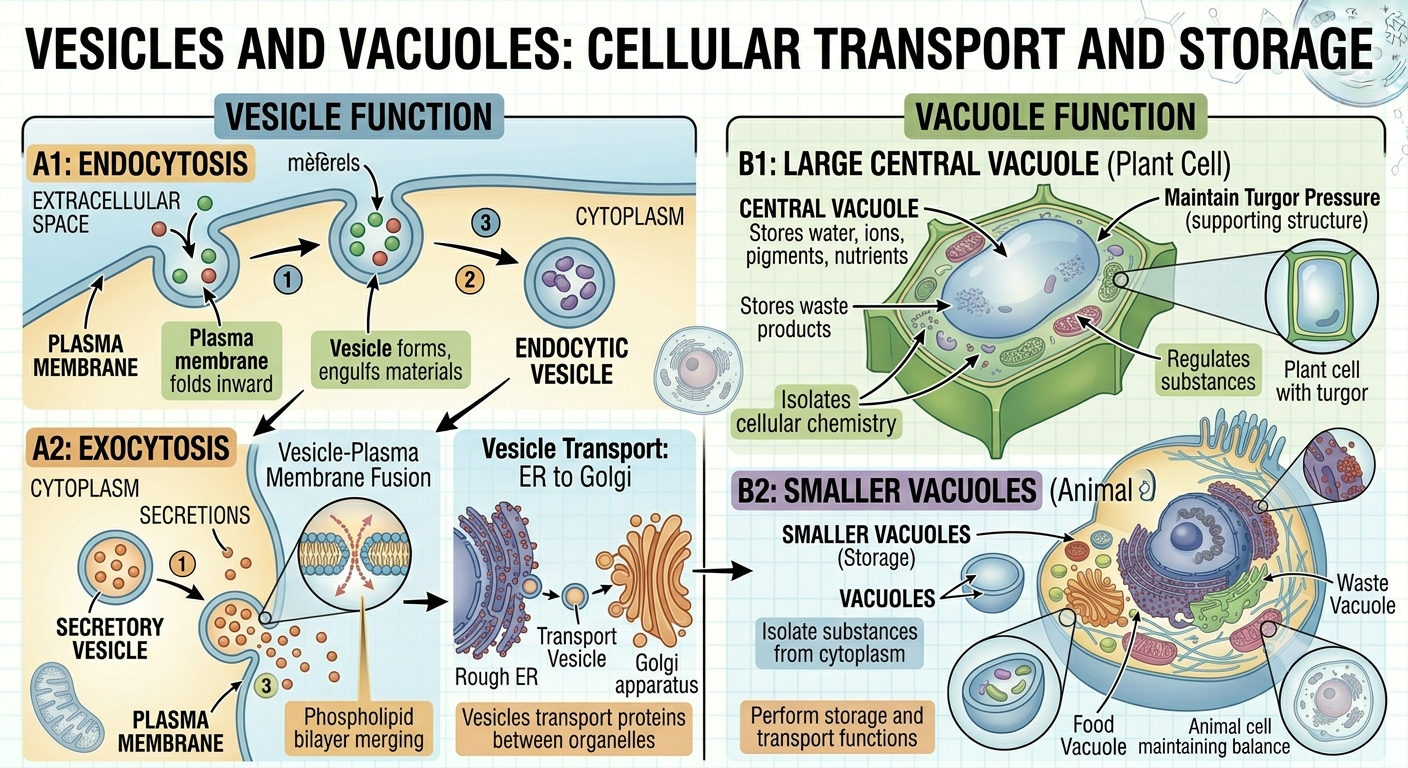
Figure 12. Vesicles and Vacuoles. Vesicles act as essential transport vehicles for moving substances between organelles and across the plasma membrane via endocytosis and exocytosis. Meanwhile, vacuoles serve as specialized storage compartments that regulate internal cellular chemistry, support plant structure through turgor pressure, and manage waste products.
Lysosomes
Lysosomes are membrane-bound organelles containing digestive enzymes capable of breaking down biomolecules such as proteins, carbohydrates, lipids, and nucleic acids. These organelles function as the recycling centers of the cell. Lysosomes digest food particles brought into the cell through endocytosis and also break down damaged organelles in a process called autophagy. The recycled molecular components can then be reused to build new cellular structures. Lysosomes also contribute to programmed cell death during development, allowing organisms to shape tissues and remove unnecessary cells.
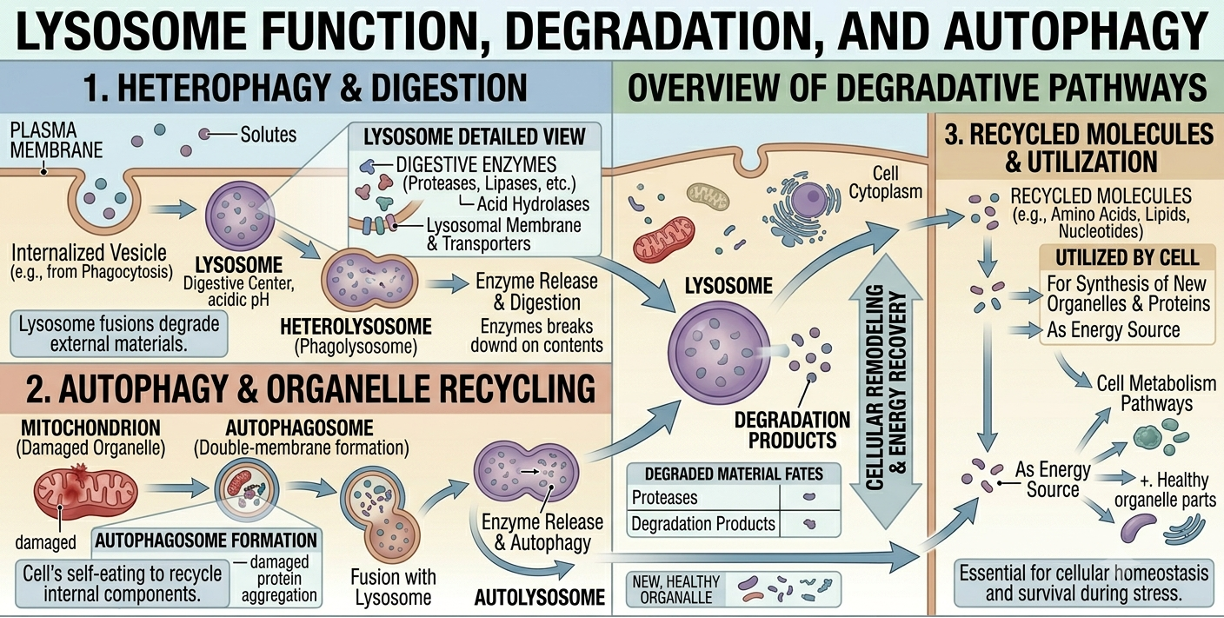
Figure 13. Lysosomal digestion and recycling. Lysosomes contain digestive enzymes that break down engulfed materials and damaged organelles. The resulting molecular components can be recycled and reused by the cell.
The Cytoskeleton
The cytoskeleton is a network of protein filaments extending throughout the cytoplasm. It provides structural support, maintains cell shape, and helps organize organelles within the cell. The cytoskeleton also plays critical roles in movement, intracellular transport, and cell division. Some cytoskeletal structures extend beyond the cell surface to form cilia and flagella, which enable movement of cells or movement of fluids across cell surfaces. In eukaryotic cells, motor proteins move vesicles and organelles along cytoskeletal tracks, allowing efficient transport throughout the cell.
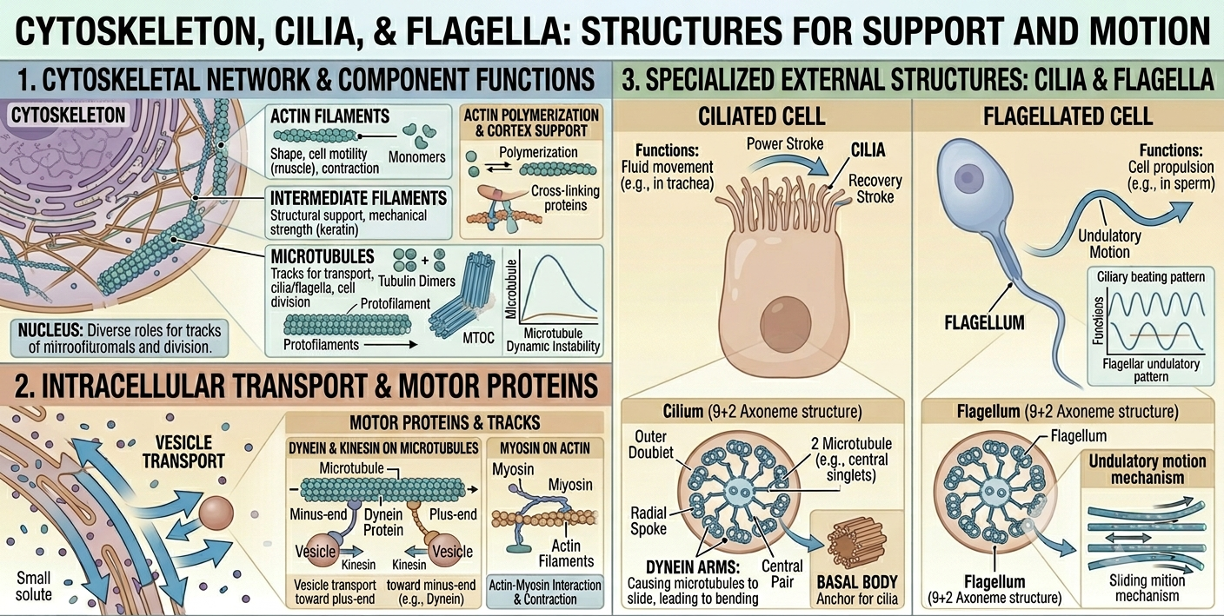
Figure 14. Functions of the cytoskeleton. The cytoskeleton maintains cell shape, organizes organelles, transports vesicles, and enables cellular movement through structures such as cilia and flagella.
Mitochondria
Mitochondria are organelles responsible for most ATP production in eukaryotic cells through cellular respiration. They act as the cell’s main energy-converting structures, breaking down nutrients like glucose and using the released energy to produce ATP, the cell’s primary usable energy source. They are surrounded by a double membrane: an outer membrane and a highly folded inner membrane. The outer membrane serves as a protective boundary and controls movement of small molecules into and out of the organelle. The inner membrane is more specialized and contains proteins involved in ATP production and the electron transport chain. Its folds, called cristae, greatly increase surface area, allowing more space for energy-generating reactions to occur efficiently. Inside the inner membrane is the matrix, a dense fluid containing enzymes needed for key stages of cellular respiration, especially the citric acid cycle. The matrix also contains mitochondrial ribosomes and mitochondrial DNA, allowing mitochondria to produce some of their own proteins and replicate independently of the cell nucleus.
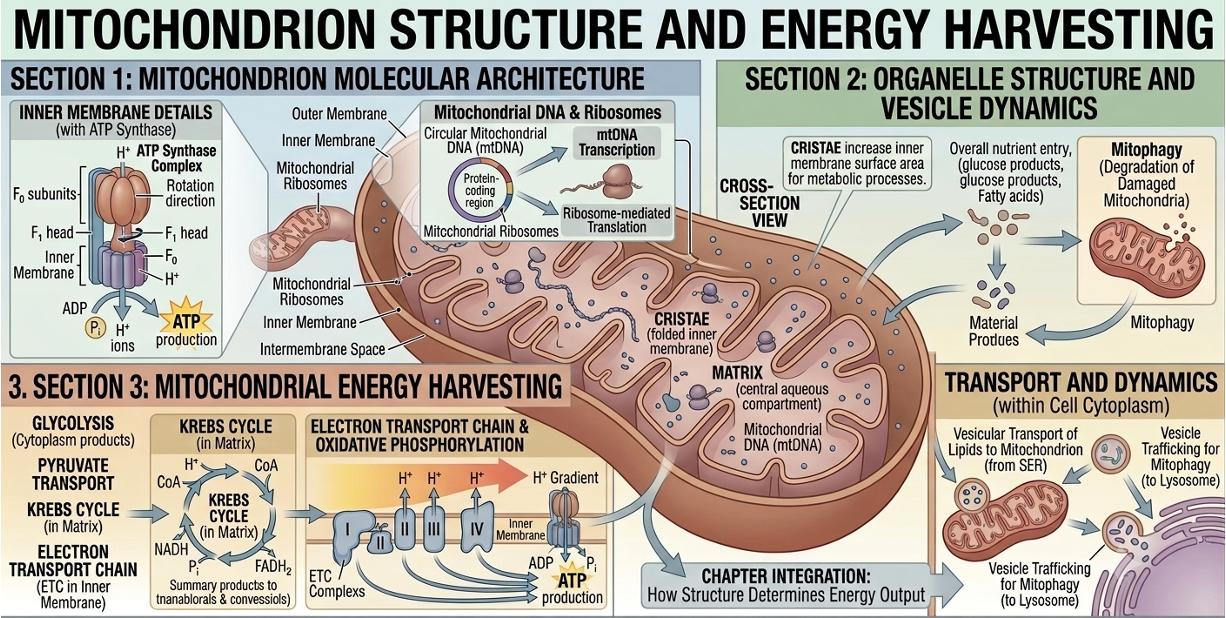
Figure 15. Structure of a mitochondrion. Mitochondria produce most ATP in eukaryotic cells through cellular respiration. Their folded inner membranes increase surface area for ATP-generating reactions.
Chloroplasts
Chloroplasts are photosynthetic organelles found in plants and algae. They contain the pigment chlorophyll, which captures light energy during photosynthesis. Chlorophyll is embedded within membranes called thylakoids, which are arranged into stacks known as grana. During photosynthesis, chloroplasts convert light energy into chemical energy stored in glucose molecules. Like mitochondria, chloroplasts possess their own DNA and ribosomes and are believed to have originated through endosymbiosis, specifically from ancient photosynthetic bacteria called cyanobacteria.
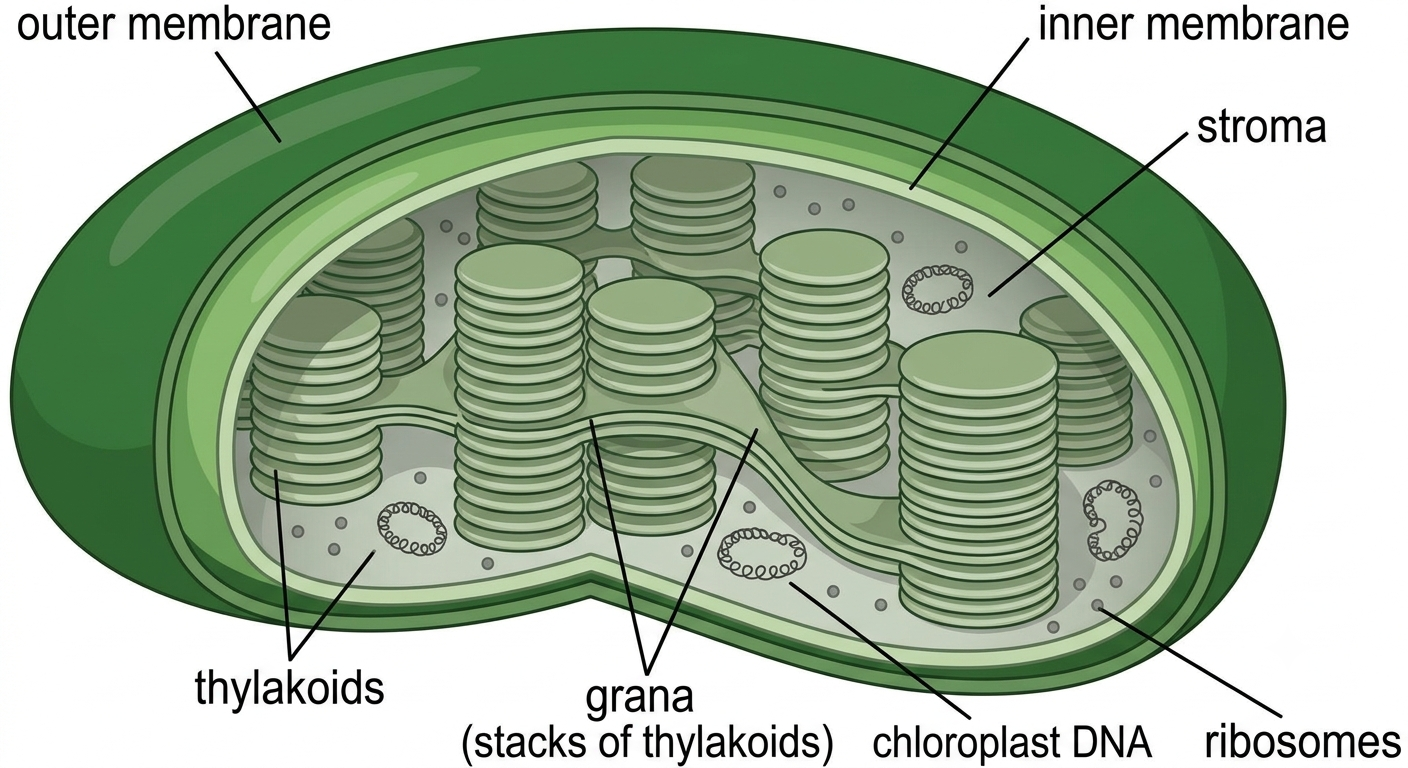
Figure 16. Structure of a chloroplast. Chloroplasts carry out photosynthesis in plant and algal cells. Light-dependent reactions occur on the thylakoid membranes, producing ATP and NADPH, while the stroma is the site of sugar formation during the Calvin cycle.
Mitochondria and Chloroplast’s Bacterial Origin
Mitochondria are believed to have evolved from ancient bacteria through endosymbiosis, where an early eukaryotic cell engulfed an aerobic bacterium that was not digested but instead formed a long-term symbiotic relationship. Over time, this bacterium became integrated into the host cell and evolved into the modern mitochondrion. Evidence for this includes the fact that mitochondria contain their own circular DNA, similar to bacterial genomes. They also have bacterial-like ribosomes, allowing them to produce some proteins independently. In addition, mitochondria have a double membrane, consistent with one membrane coming from the host cell and the other from the original bacterium. Together, these features support the endosymbiotic theory and explain why mitochondria retain bacterial characteristics despite being essential eukaryotic organelles.
Chloroplasts are believed to have originated through endosymbiosis, a process in which a eukaryotic host cell engulfed a free-living photosynthetic bacterium, most likely a cyanobacterium, without digesting it. Instead of being broken down, the internalized bacterium formed a mutually beneficial relationship with the host cell: it provided the host with sugars produced through photosynthesis, while the host supplied protection and access to nutrients. Over evolutionary time, this engulfed bacterium became permanently integrated into the host cell, gradually losing many of its independent genes as some were transferred to the host nucleus. Evidence for this origin includes chloroplasts having their own circular DNA, bacterial-like ribosomes, and a double membrane consistent with engulfment.
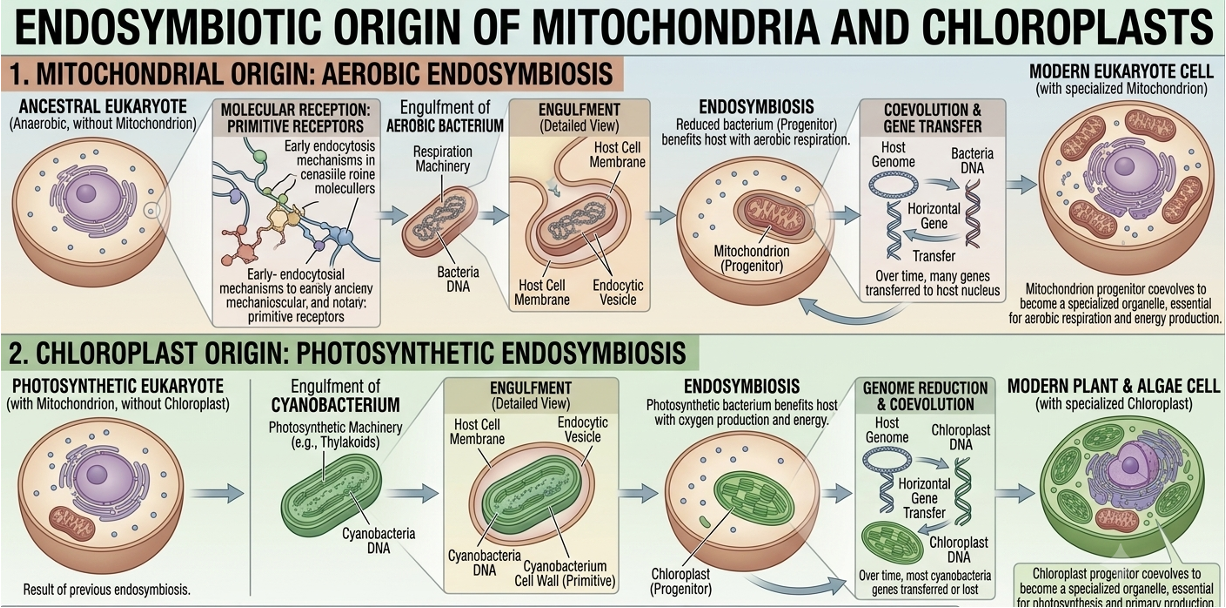
Figure 17. Endosymbiotic origin of mitochondria and chloroplasts. Mitochondria and chloroplasts likely evolved from free-living bacteria engulfed by ancestral eukaryotic cells. Evidence includes their circular DNA, bacterial-like ribosomes, and double membranes.
Plant Cells vs. Animal Cells
Plant and animal cells are both eukaryotic and share many organelles, including nuclei, mitochondria, ribosomes, and endoplasmic reticulum. However, plant cells possess several structures not found in animal cells. Plant cells contain chloroplasts for photosynthesis, a large central vacuole for water storage and support, and a rigid cell wall composed primarily of cellulose. These structures give plant cells a more rigid and rectangular shape. Animal cells lack cell walls and chloroplasts and generally display a more flexible and irregular shape. Despite these differences, both plant and animal cells rely on the same fundamental cellular processes to sustain life.
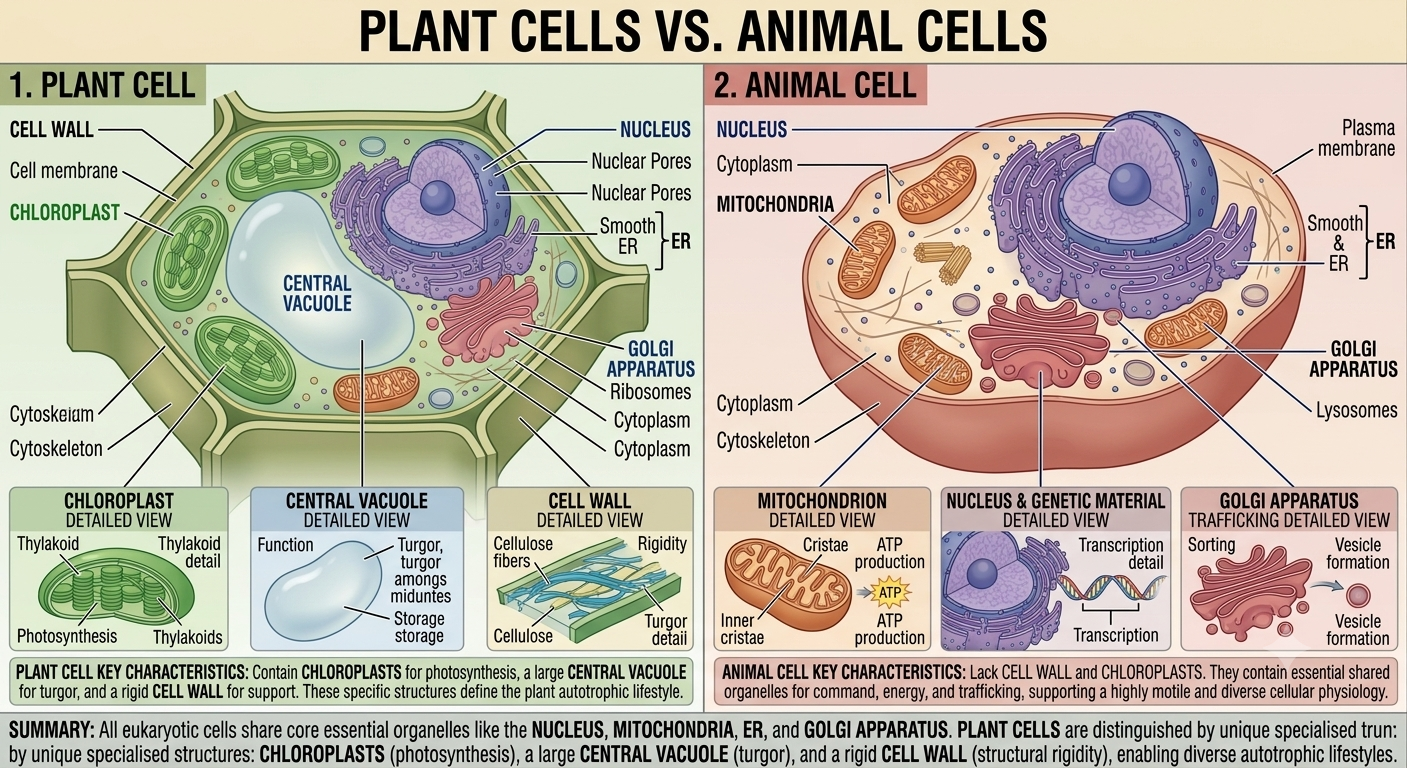
Figure 18. Comparison of plant and animal cells. Plant and animal cells share many eukaryotic organelles but differ in several important ways. Plant cells possess chloroplasts, a large central vacuole, and a cellulose cell wall, whereas animal cells lack these structures and are generally more flexible in shape.