
Chapter: Speciation
Speciation and Gene Flow
Speciation is the evolutionary process through which new species arise. It is one of the major ways biodiversity increases over time. Although speciation can occur through several pathways, most cases begin when gene flow between populations is reduced. Gene flow is the movement of alleles between populations through mating, migration, or other forms of genetic exchange. When gene flow is high, populations tend to remain genetically similar. When gene flow is reduced or interrupted, populations can begin to diverge through mutation, natural selection, genetic drift, and sexual selection. Speciation does not always begin with complete isolation. In many cases, gene flow declines gradually. Populations may still exchange some genes while becoming increasingly different from one another. For this reason, speciation is often described as a continuum. At one end are populations that freely interbreed. In the middle are populations that differ genetically or ecologically but may still produce some hybrids. At the far end are fully separate species that no longer exchange genes under natural conditions. A useful example of reduced gene flow leading to speciation is the geographic separation of South America and Africa. These continents were once connected as part of a larger landmass. As plate tectonic movement opened the Atlantic Ocean, populations on opposite sides became separated by a growing ocean barrier. Over millions of years, isolation allowed many lineages on each continent to evolve independently. Geographic separation does not automatically produce new species, but it creates the conditions in which speciation can occur.
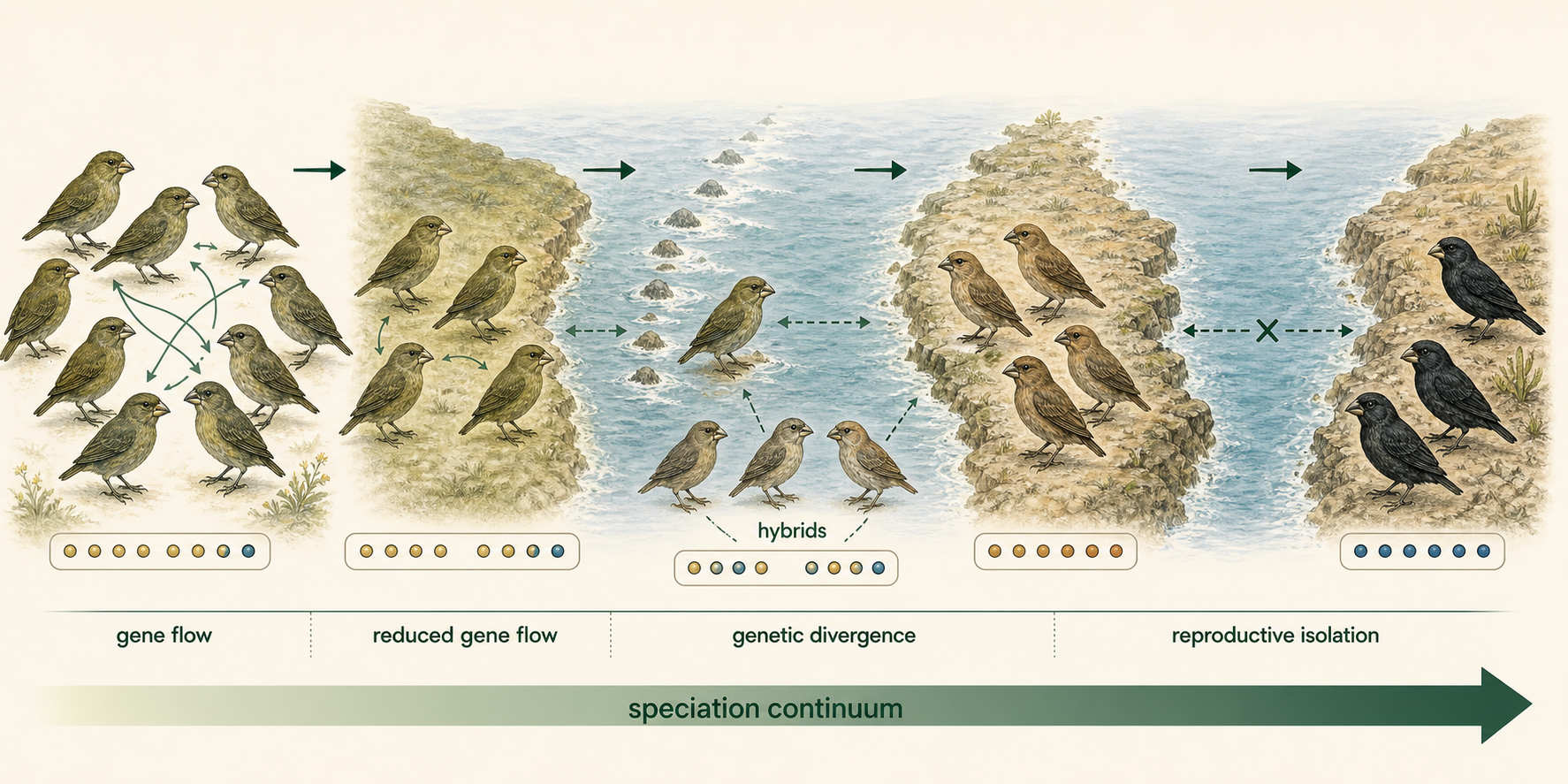
Figure 1. Speciation and Gene Flow. Speciation often begins when gene flow between populations is reduced. As populations exchange fewer alleles, mutation, natural selection, genetic drift, and sexual selection can cause them to diverge over time.
Reproductive Isolation
For speciation to be completed, populations must become reproductively isolated. Reproductive isolation occurs when barriers prevent populations from successfully exchanging genes. These barriers may prevent mating or fertilization, or they may reduce the survival or fertility of hybrid offspring. Reproductive barriers are usually grouped into two major categories: prezygotic barriers and postzygotic barriers.
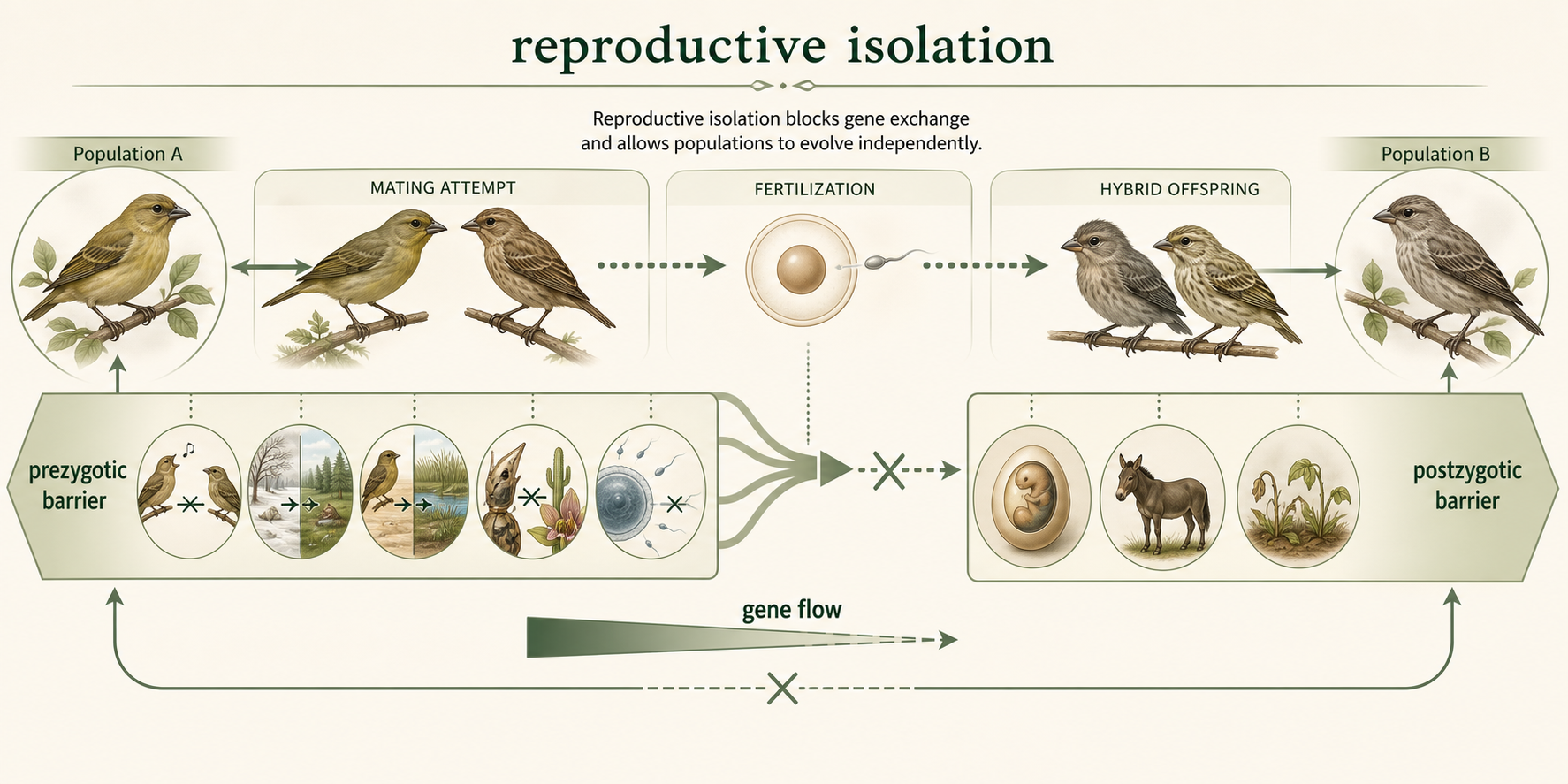
Figure 2. Reproductive Isolation. Reproductive isolation occurs when barriers prevent populations from successfully exchanging genes. These barriers may act before fertilization or after fertilization by reducing hybrid survival or fertility.
Reproductive Isolation: Prezygotic barriers
Prezygotic barriers act before a zygote forms. These barriers prevent members of different populations or species from mating or from successfully fertilizing an egg. Habitat isolation occurs when organisms live in the same general region but use different habitats. For example, two species of garter snakes may live in the same area, but one species lives mainly in water while the other lives mainly on land. Because they rarely encounter each other for mating, gene flow is reduced. Temporal isolation occurs when organisms reproduce at different times of day, seasons, or years. For example, two closely related frog species may live in the same pond, but one breeds in early spring while the other breeds in late summer. Because their breeding seasons do not overlap, they are unlikely to mate. Behavioral isolation occurs when mating signals, songs, scents, dances, or courtship behaviors differ. For example, many bird species recognize mates by specific songs. If two closely related bird species have different songs, individuals may only respond to the song of their own species. Mechanical isolation occurs when reproductive structures are incompatible. For example, some closely related insect species have reproductive body parts that do not fit together properly, preventing successful mating. Gametic isolation occurs when sperm and egg cells cannot successfully fuse. For example, many marine animals release sperm and eggs into the water, but chemical signals on the surface of the egg often allow fertilization only by sperm from the same species.
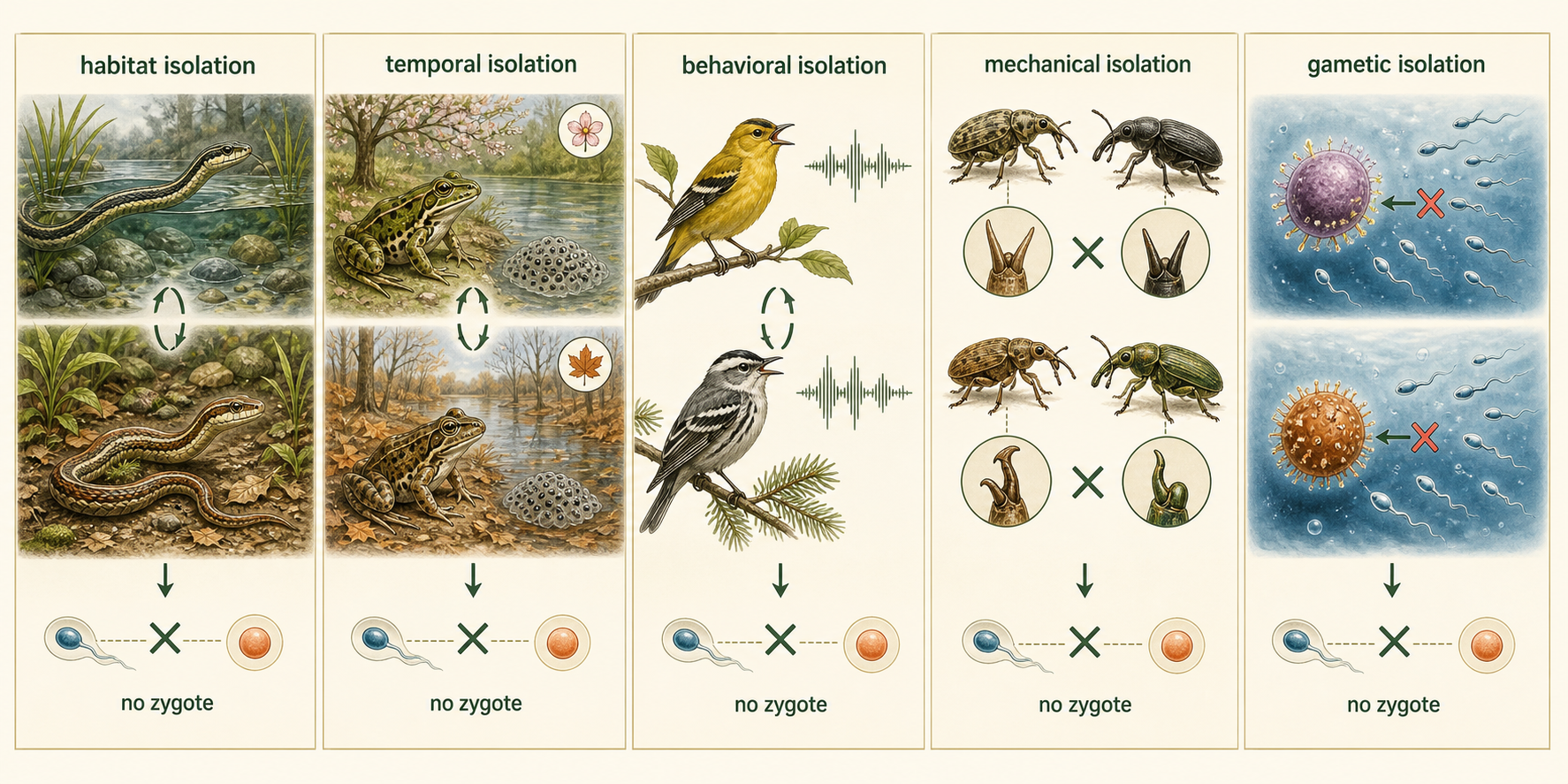
Figure 3. Prezygotic Barriers. Prezygotic barriers act before a zygote forms. They prevent mating or fertilization through differences in habitat, timing, behavior, reproductive structures, or gamete compatibility.
Reproductive Isolation: Postzygotic barriers
Postzygotic barriers act after fertilization. These barriers reduce the survival or fertility of hybrid offspring. Hybrid inviability occurs when hybrid embryos fail to develop properly or die before reaching reproductive age. For example, some crosses between closely related frog species produce embryos that begin developing but do not survive long enough to become adults. Hybrid sterility occurs when hybrids survive but cannot produce functional gametes. For example, a horse and a donkey can produce a mule, but mules are usually sterile because their chromosomes do not pair properly during meiosis. Hybrid breakdown occurs when first-generation hybrids are viable and fertile, but later generations are weak, sterile, or less fit. For example, two closely related plant species may produce healthy first-generation hybrids, but when those hybrids reproduce, their offspring may have reduced fertility or poor survival.Horses, Donkeys, and Mules Horses and donkeys provide a familiar example of postzygotic isolation. A horse and a donkey can mate and produce a mule, but mules are usually sterile. This does not mean fertilization is impossible; rather, it means gene flow is strongly limited because the hybrid offspring usually cannot pass genes to the next generation. This supports the classification of horses and donkeys as separate species.
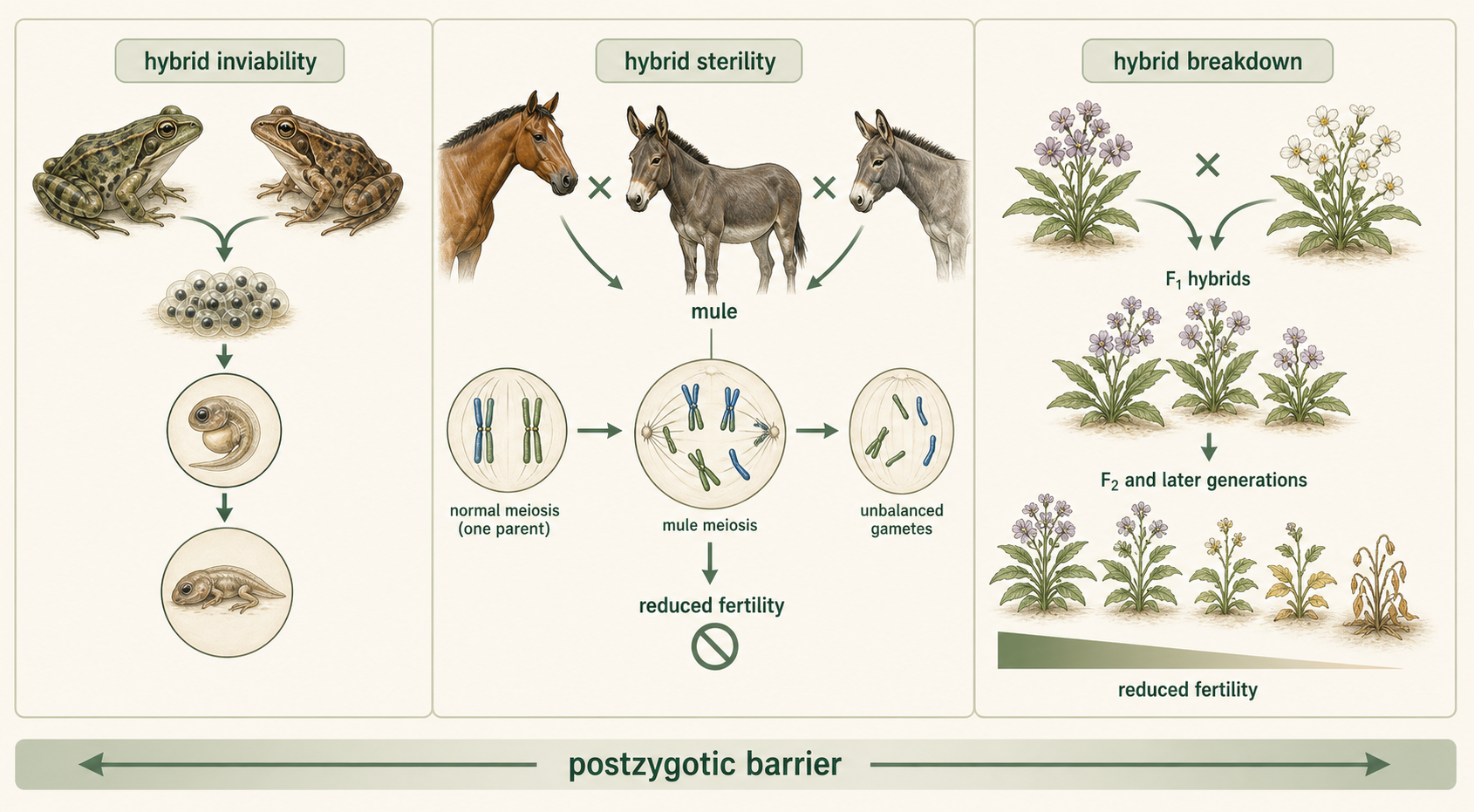
Figure 4. Postzygotic Barriers. Postzygotic barriers act after fertilization. They reduce gene flow when hybrid offspring fail to survive, cannot reproduce, or produce later generations with reduced fitness.
What Is a Species?
To understand speciation, we must first ask a deceptively difficult question: What is a species? Biologists have proposed several ways to define species. No single definition works perfectly for all organisms, because life is too varied. Some organisms reproduce sexually, some reproduce asexually, some hybridize, some are known only from fossils, and some are nearly identical in appearance despite being genetically distinct. For this reason, species concepts are tools. Each one emphasizes a different kind of evidence. Next, we will explore three different ways to define a species: 1) Biological Species Concept, 2) Morphological Species Concept, and 3) Phylogenetic Species Concept.
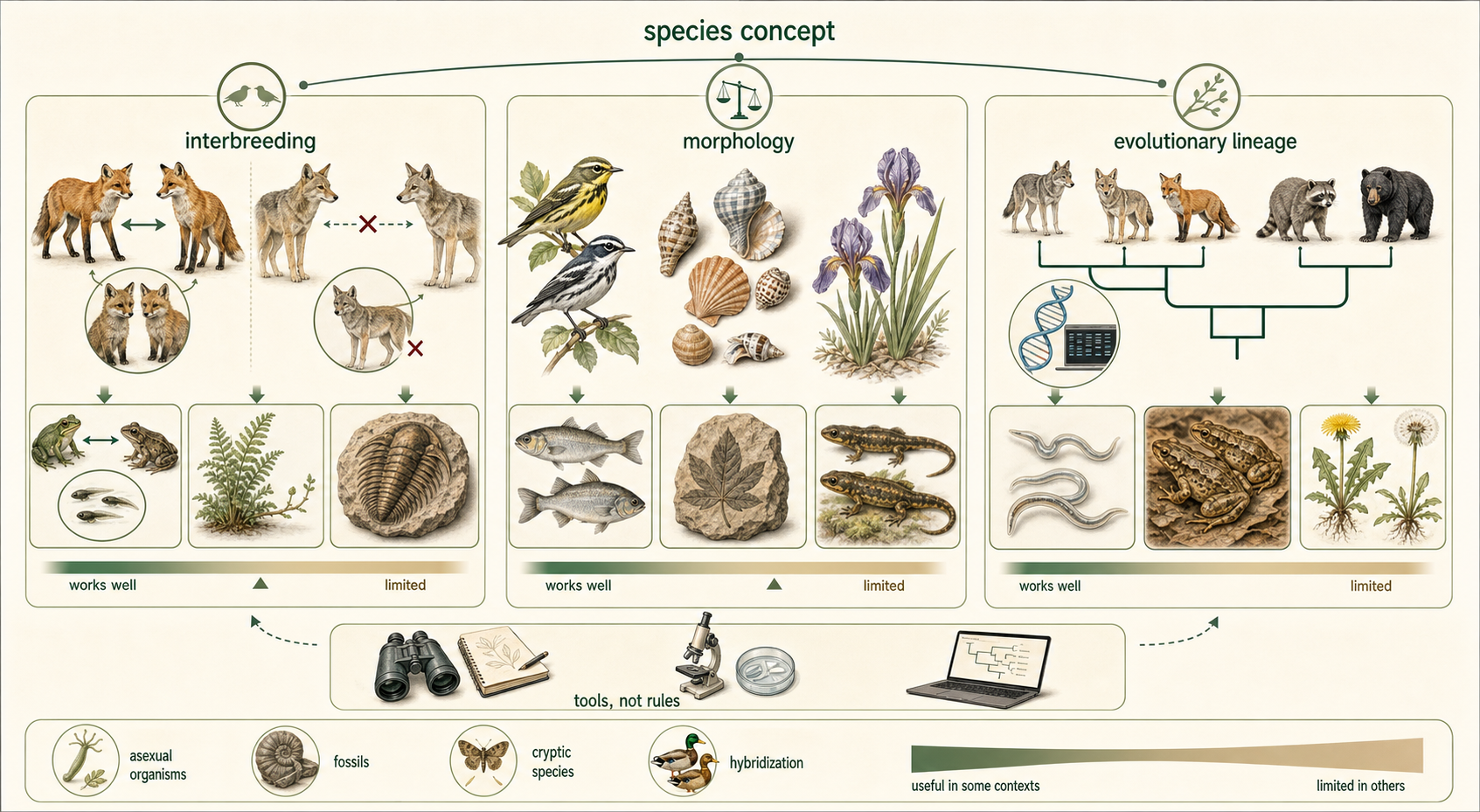
Figure 5. What Is a Species? Biologists use different species concepts because no single definition works perfectly for all organisms. Species may be defined by reproductive isolation, observable traits, or evolutionary history.
The Biological Species Concept
The biological species concept is one of the most widely used definitions of a species. It defines a species as a group of natural populations whose members can interbreed with one another and produce viable, fertile offspring, but are reproductively isolated from other such groups. In this context, “viable” means the offspring can survive, and “fertile” means the offspring can reproduce.
Strengths of the Biological Species Concept
The biological species concept is especially useful for studying sexually reproducing organisms that are alive today. It helps explain why organisms that look similar may still be separate species. For example, some salamanders, insects, or birds may appear almost identical but do not interbreed because of differences in behavior, genetics, timing, or reproductive compatibility. These are often called cryptic species because their differences are hidden or difficult to detect by appearance alone.
Limitations of the Biological Species Concept
However, the biological species concept has important limitations. It cannot be applied easily to extinct organisms because we cannot directly observe whether fossil species could interbreed. It also does not work well for organisms that reproduce asexually, such as many bacteria and some single-celled eukaryotes. In addition, it can be difficult to apply to populations that live in different geographic regions. Two populations may be capable of interbreeding if brought together, but if they never naturally meet, their species status may be uncertain. Another complication is hybridization. Some clearly distinct species can occasionally produce fertile hybrids. This is common in plants and also occurs in some animals. Wolves, coyotes, and domestic dogs, for example, can exchange genes in some circumstances. Hybridization shows that species boundaries are sometimes porous rather than absolute.
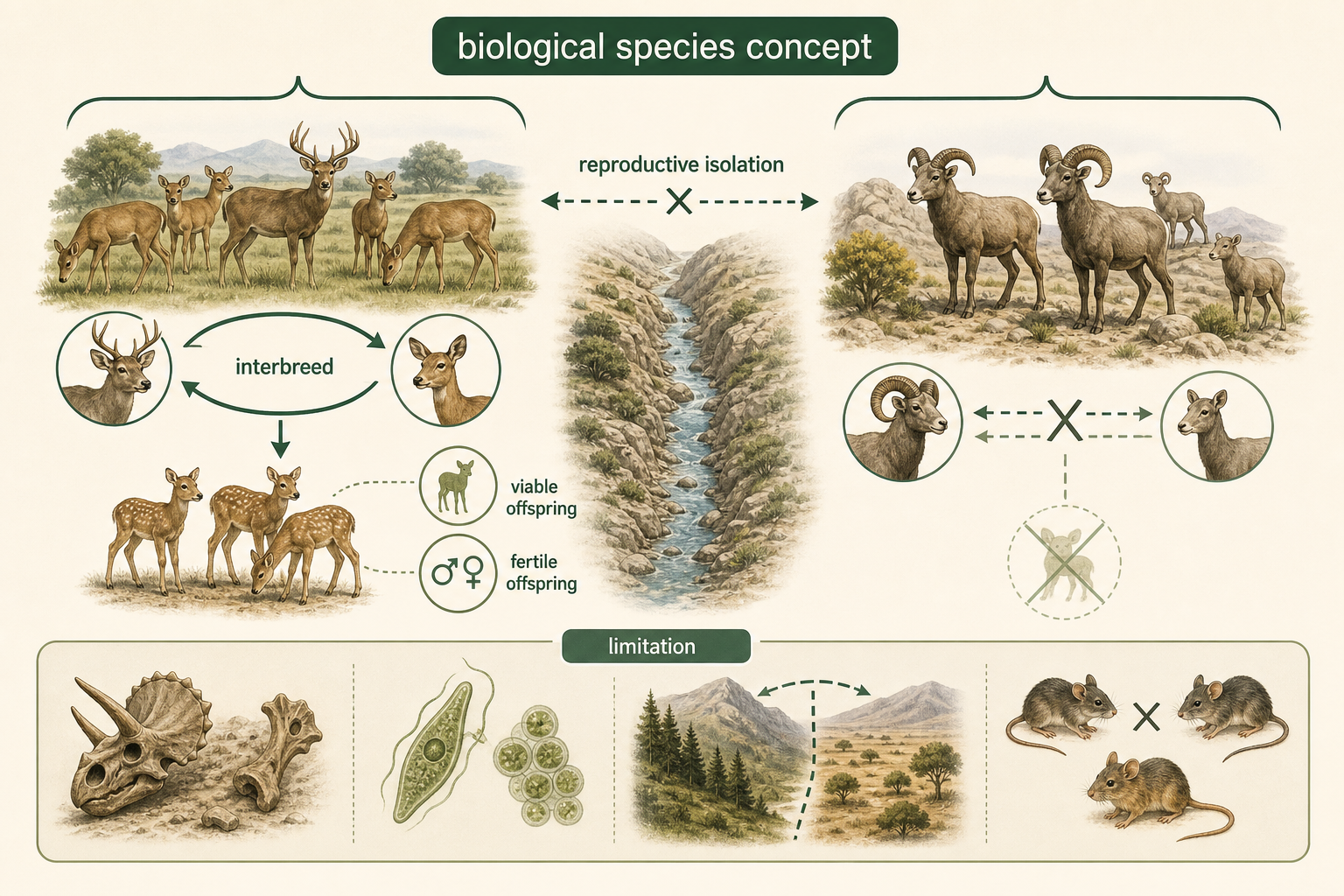
Figure 6. Biological Species Concept. The biological species concept defines a species as natural populations that can interbreed and produce viable, fertile offspring while remaining reproductively isolated from other groups.
The Morphospecies Concept
Another commonly used approach is the morphospecies concept, defining species based on observable physical traits. Under this approach, organisms are classified into different species if they differ consistently in body form, structure, size, color, anatomy, or other visible features. The morphospecies concept is widely used in field biology, paleontology, and everyday species identification. Bird watchers, plant collectors, fossil researchers, and naturalists often rely on morphology because they cannot perform breeding studies or DNA analyses every time they encounter an organism. Field guides are usually built around this concept because physical traits are often the most practical information available.
Strengths of the Morphospecies Concept
The morphospecies concept has several strengths. It can be used for fossils. It can be used for organisms that reproduce asexually. It can be applied when populations do not overlap geographically. It also allows rapid identification in the field.
Limitations of the Morphospecies Concept
However, it also has weaknesses. Some species look nearly identical but are reproductively or genetically distinct. These cryptic species are easy to miss if classification depends only on appearance. The opposite problem also occurs: individuals within the same species can look dramatically different. Domestic dogs provide a clear example. Chihuahuas and Great Danes differ greatly in size and shape, yet they belong to the same species, Canis familiaris, because they are part of the same domesticated lineage and can share genes through breeding. Morphology can also be influenced by age, sex, environment, nutrition, and development. Males and females of the same species may look different. Juveniles may look unlike adults. Individuals living in different environments may develop different traits even when they are genetically similar. Because of this, using morphology alone can sometimes split one species into too many groups or lump several species into one.
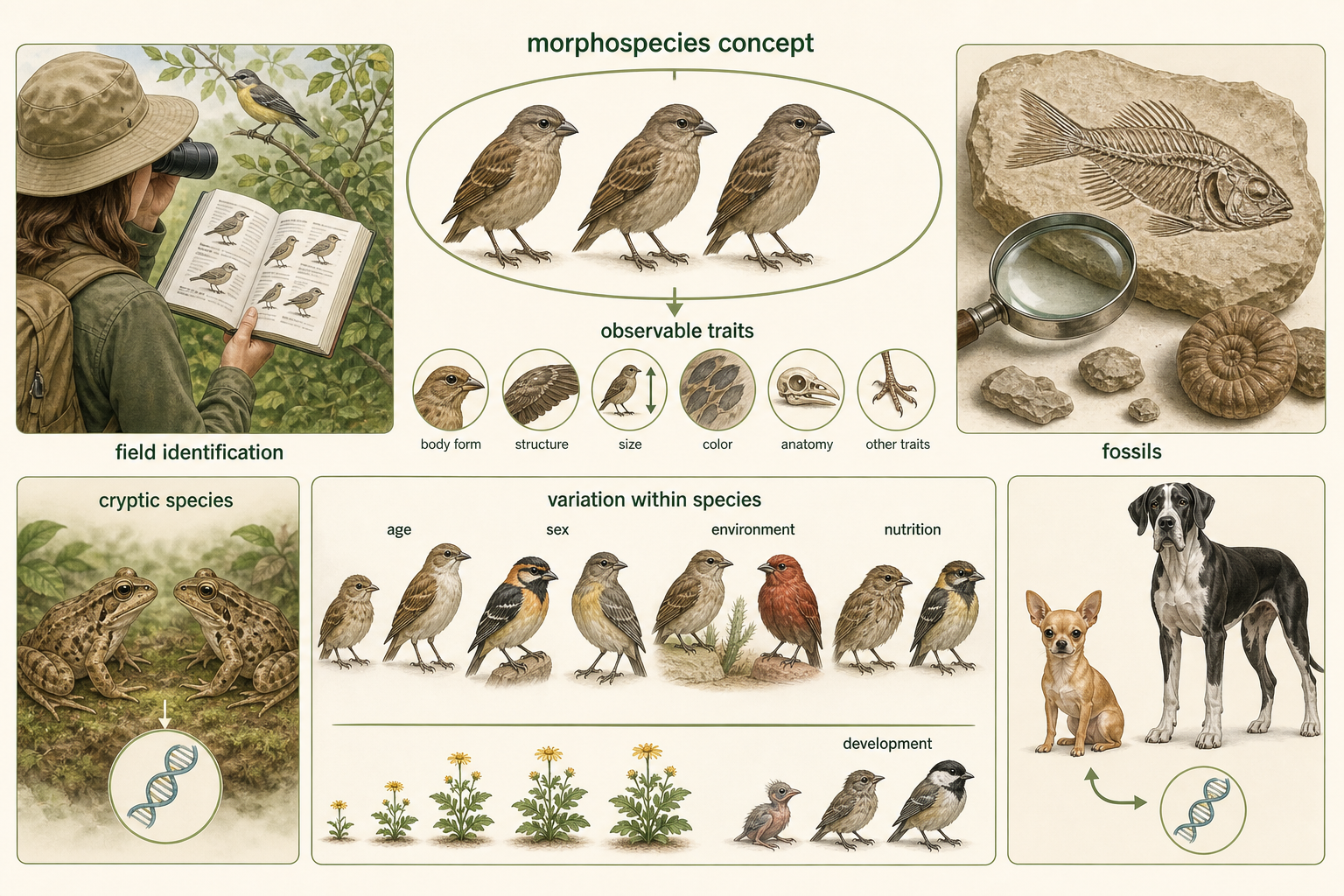
Figure 7. Morphospecies Concept. The morphospecies concept defines species using observable physical traits. It is useful in field biology and paleontology, but morphology alone can miss cryptic species or overemphasize variation within a species.
The Phylogenetic Species Concept
A third approach is the phylogenetic species concept. This concept defines species based on evolutionary history. A species is often treated as the smallest distinct evolutionary lineage that shares a common ancestry and can be distinguished from other lineages by unique inherited traits. These traits may be physical, behavioral, ecological, or genetic. The phylogenetic species concept became especially powerful with advances in DNA sequencing. Genetic data allow scientists to compare organisms at the molecular level and reconstruct their evolutionary relationships. A group that includes an ancestor and all of its descendants is called a monophyletic group, or clade. Shared derived traits that help identify a clade are called synapomorphies. For example, feathers are a synapomorphy of birds, and mammary glands are a synapomorphy of mammals.
Strengths of the Phylogenetic Species Concept
The phylogenetic species concept has several advantages. It can identify cryptic species that look alike but have distinct evolutionary histories. It does not require direct observation of mating. It can be used with living organisms, fossils, and asexual lineages, as long as enough evidence is available. It also reflects the branching pattern of evolution more directly than definitions based only on appearance or breeding. A well-known example involving phylogenetic evidence is the distinction between African savanna elephants and African forest elephants. For many years, African elephants were commonly treated as one species with different forms or subspecies. Genetic and morphological evidence now supports recognizing two African elephant species: the larger savanna elephant, Loxodonta africana, and the smaller forest elephant, Loxodonta cyclotis. This example shows how DNA evidence can reveal deep evolutionary separation that may not be obvious from a simple field observation.
Limitations of the Phylogenetic Species Concept
However, the phylogenetic species concept also has limitations. It can greatly increase the number of recognized species because even small, diagnosable genetic differences may be interpreted as evidence of separate lineages. This can make field identification difficult. It also depends on the quality of the available data. Many groups of organisms remain poorly studied, and incomplete sampling can make evolutionary relationships uncertain. In addition, gene trees do not always perfectly match species histories because of hybridization, incomplete lineage sorting, or horizontal gene transfer.
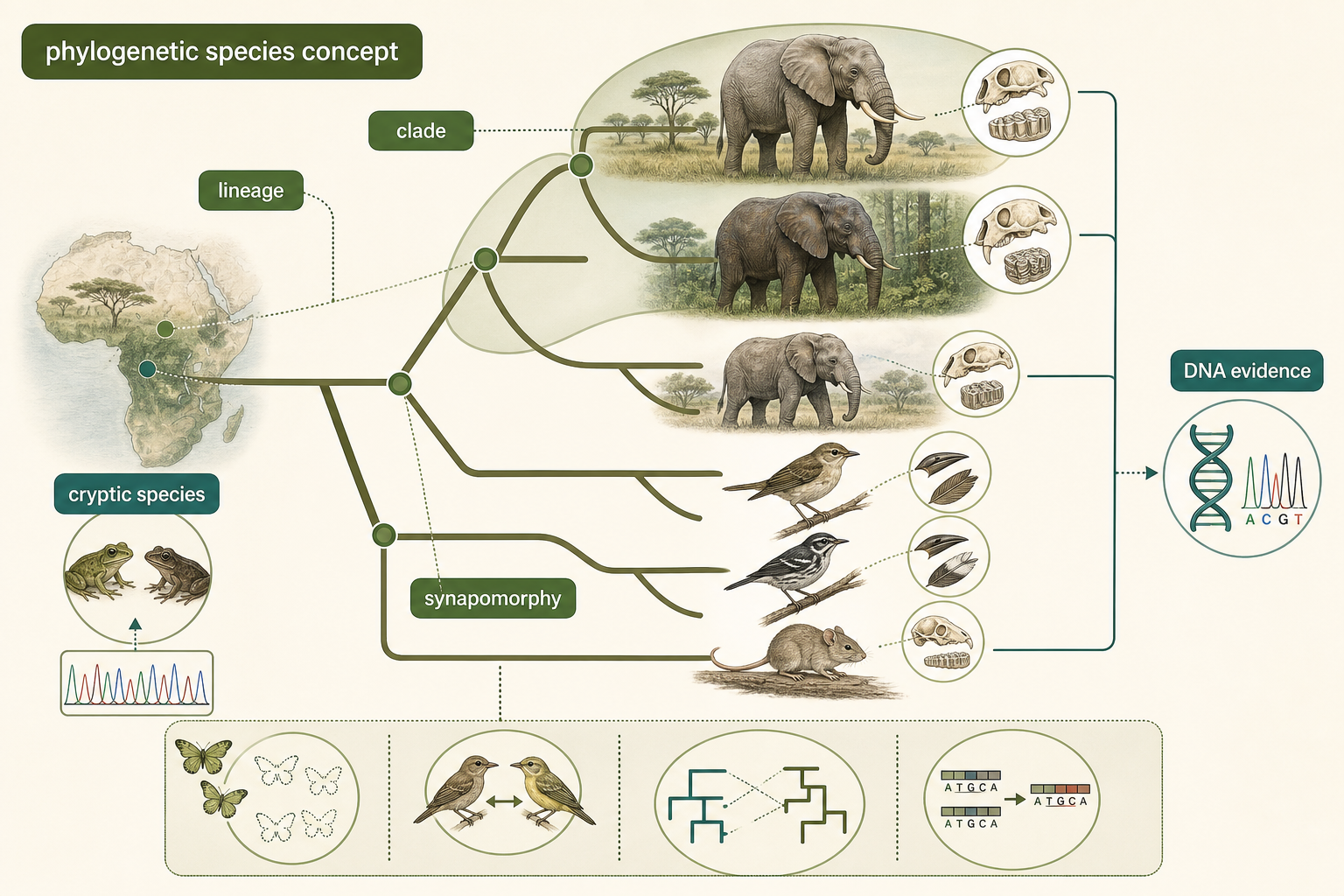
Figure 8. Phylogenetic Species Concept. The phylogenetic species concept defines species as distinct evolutionary lineages that share common ancestry and unique inherited traits. DNA evidence can reveal lineages that are difficult to recognize by appearance alone.
Subspecies
Once species are defined, the idea of subspecies becomes easier to understand. A subspecies is a geographically or genetically distinct population within a species. Subspecies often have recognizable traits and limited gene flow with other populations of the same species, but they are not usually considered fully separate species. Subspecies may represent early stages of divergence. Over time, if isolation continues and reproductive barriers strengthen, subspecies may eventually become separate species.
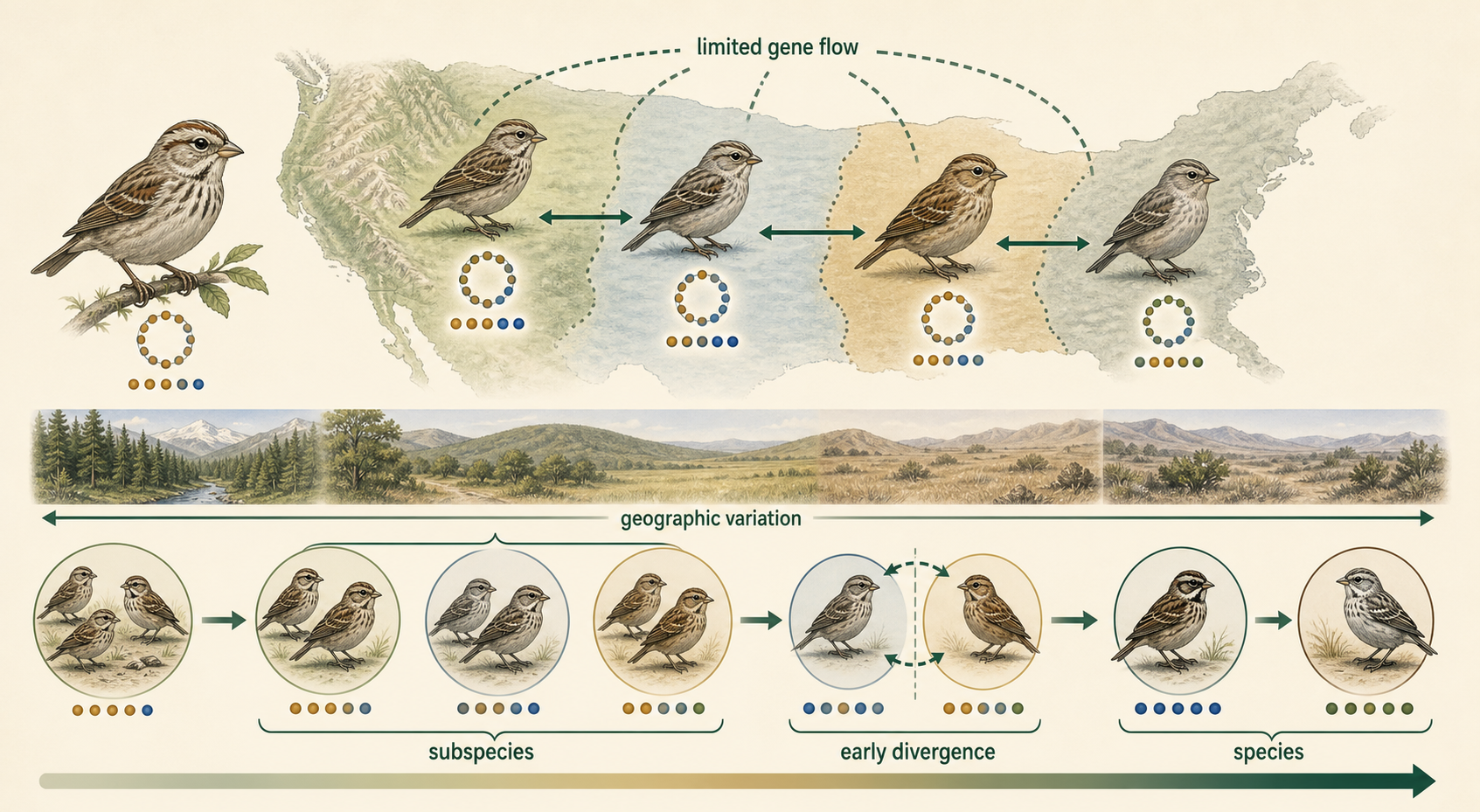
Figure 9. Subspecies. A subspecies is a geographically or genetically distinct population within a species. Subspecies often have recognizable traits and limited gene flow, but they are not usually considered fully separate species.
Allopatric Speciation
Speciation often begins with genetic isolation caused by physical separation. This is called allopatric speciation. In allopatric speciation, populations become separated by geography. Once separated, they no longer exchange genes freely. Different mutations arise in each population. Genetic drift may change allele frequencies, especially in small populations. Natural selection may favor different traits in different environments. Over many generations, the populations may become so different that they can no longer interbreed successfully, even if they later come back into contact.
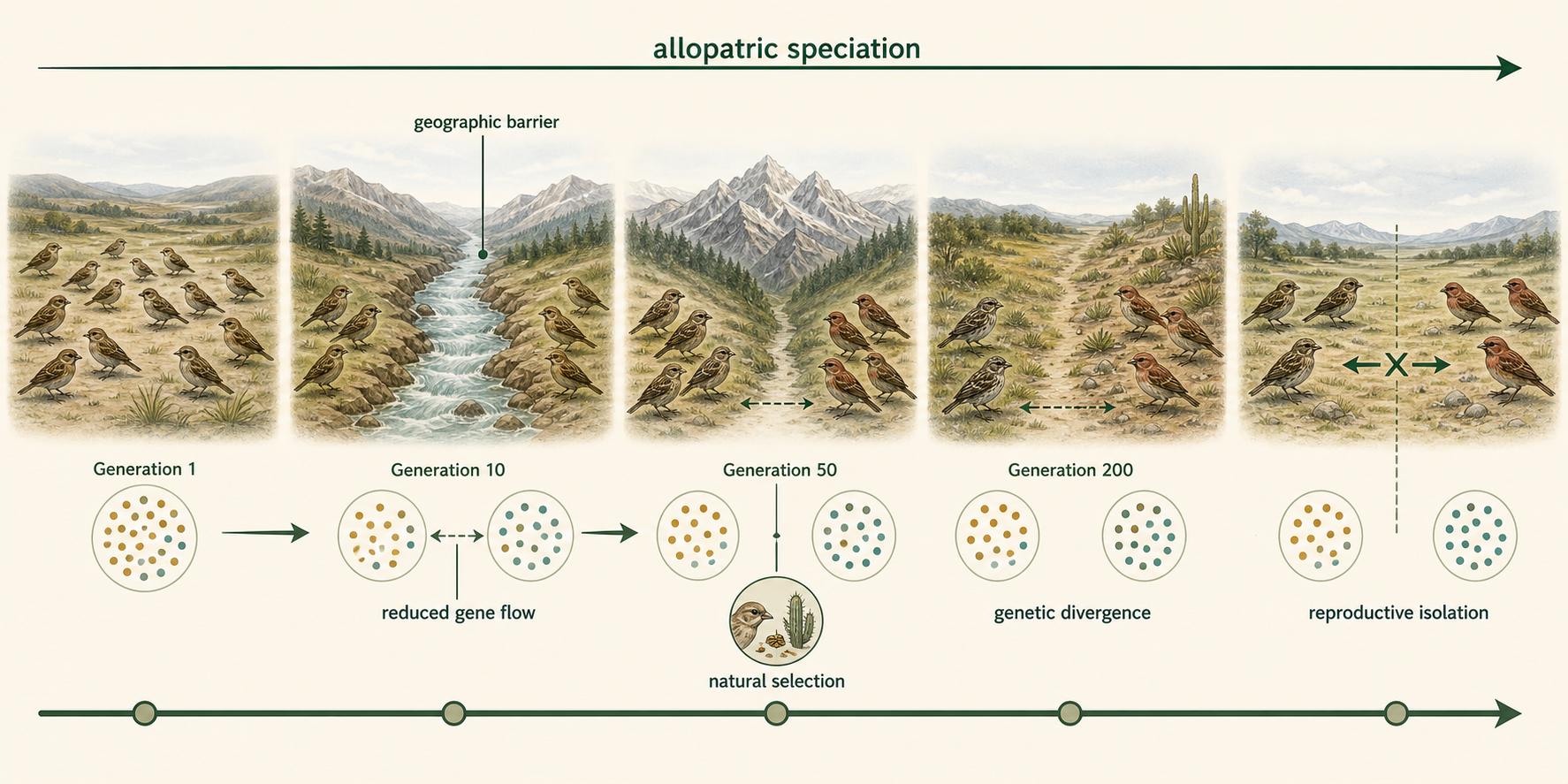
Figure 10. Allopatric Speciation. Allopatric speciation begins when populations are geographically separated and no longer exchange genes freely. Over time, mutation, drift, and selection can cause divergence and reproductive isolation.
Allopatric Speciation: Dispersal
Physical separation can occur through dispersal or vicariance. Dispersal occurs when some individuals move away from the original population and colonize a new area. This may happen when organisms reach an island, cross a mountain range, move into a new lake, or become isolated at the edge of a species’ range. If the founding group is small, it may carry only a portion of the original population’s genetic diversity. This is called the founder effect. The founder effect can cause rapid genetic differences between the new population and the original population.
The Galápagos finches provide a classic example of dispersal and founder effects. The ancestors of these finches likely arrived on the Galápagos Islands from mainland South America. Once a small founding population reached the islands, its descendants spread to different islands and encountered different environments, food sources, and ecological conditions. Because each island population was partly isolated from the others, gene flow was reduced. Over time, natural selection favored different traits in different environments, especially differences in beak size and shape. For example, finches that fed mainly on large, hard seeds benefited from deeper, stronger beaks, while finches that fed on small seeds, insects, or cactus flowers benefited from different beak shapes. As these island populations continued to diverge, differences in feeding behavior, body size, song, and mating patterns helped reduce gene flow even further. Over many generations, this process contributed to the evolution of multiple finch species from a common ancestor. The Galápagos finches therefore illustrate how dispersal to new habitats, founder effects, geographic isolation, and natural selection can work together to promote speciation.
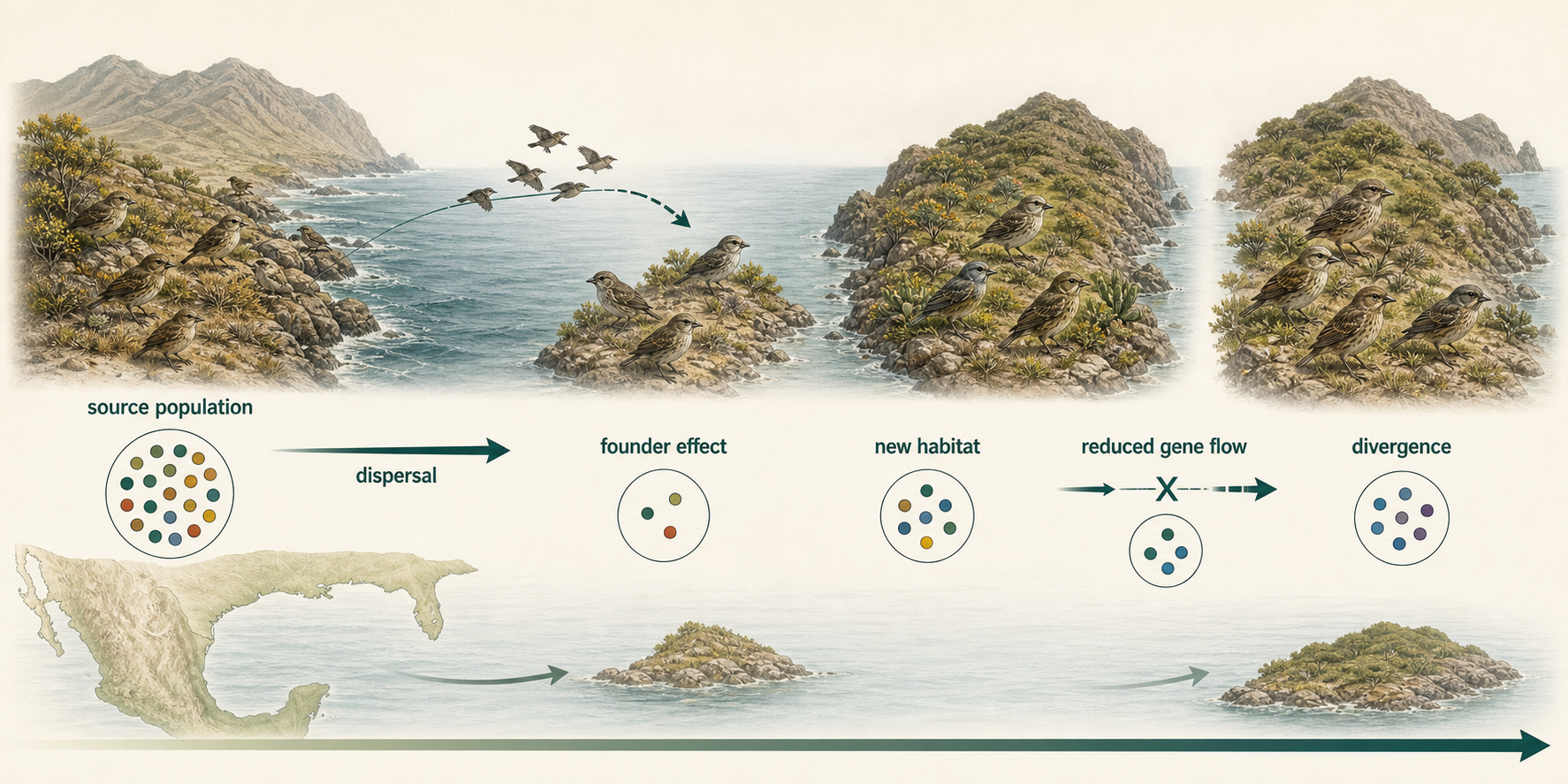
Figure 11. Allopatric Speciation by Dispersal. Dispersal can begin allopatric speciation when a small group colonizes a new area and becomes isolated. Founder effects, reduced gene flow, drift, and selection can then promote divergence.
Allopatric Speciation: Vicariance
Vicariance occurs when a barrier forms and splits an existing population. Barriers may include rivers, glaciers, mountains, deserts, canyons, lava flows, or oceans. Climate change can also create barriers by fragmenting habitats. For example, a once-continuous forest may shrink into isolated patches, separating populations that previously exchanged genes. The Grand Canyon is often used to illustrate geographic isolation, but it should be used carefully. The Kaibab squirrel on the North Rim and Abert’s squirrel populations on the South Rim are commonly discussed as examples of divergence associated with geographic and habitat isolation. However, the Kaibab squirrel is usually classified as a subspecies of Abert’s squirrel rather than a fully separate species. The example is still useful because it shows how isolation can produce recognizable population differences, but it should not be presented as a simple completed speciation event.
Sympatric Speciation
In contrast to allopatric speciation, sympatric speciation occurs without geographic separation. In sympatric speciation, populations diverge while living in the same general area. This may seem unlikely because gene flow should blend the populations together. However, sympatric speciation can occur when gene flow is reduced by ecological differences, mating preferences, behavior, or chromosome changes.
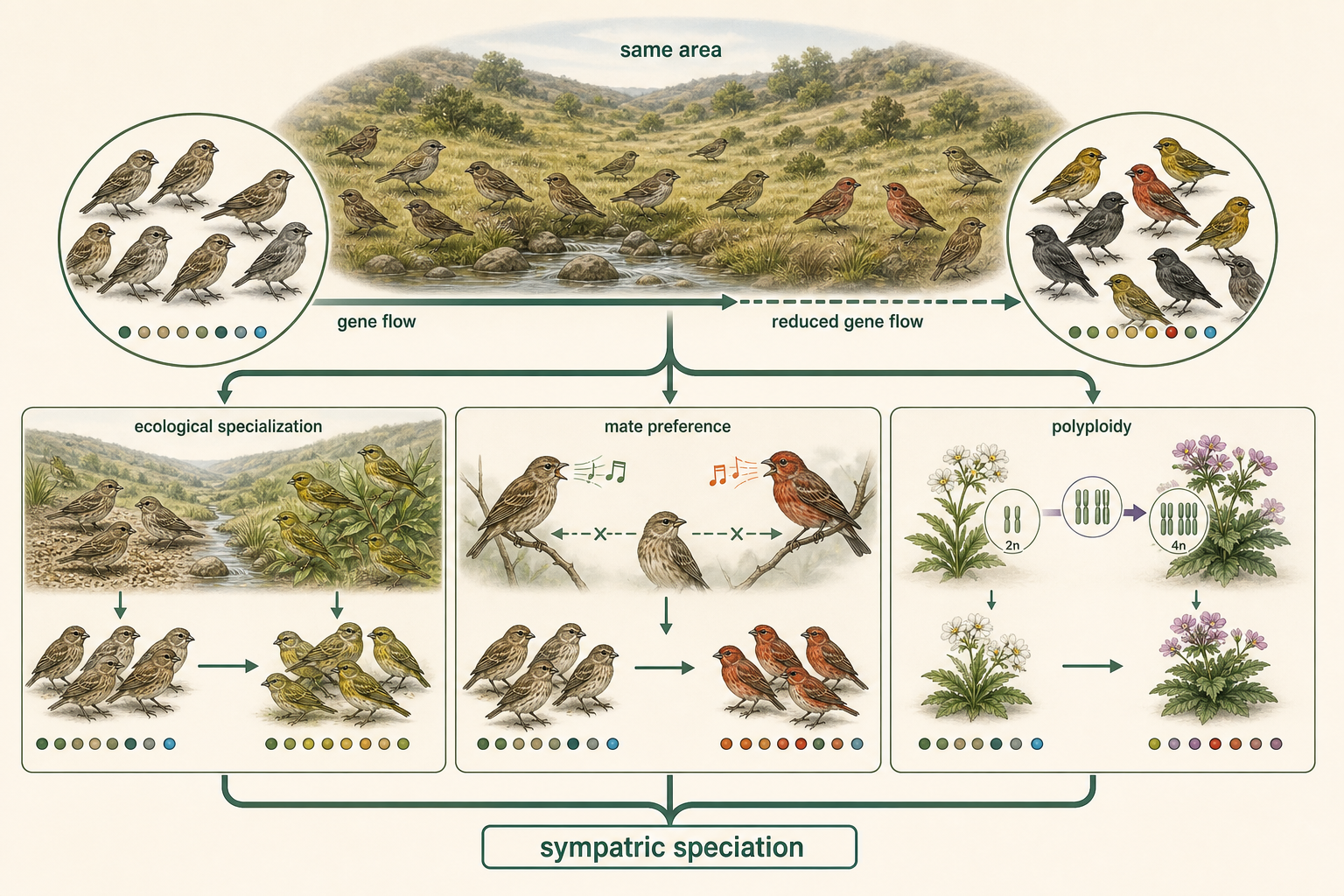
Figure 12. Sympatric Speciation. Sympatric speciation occurs when populations diverge while living in the same general area. Gene flow may be reduced by ecological differences, mating preferences, behavior, or chromosome changes.
Sympatric Speciation: Ecological Specialization
One pathway to sympatric speciation is ecological specialization. If individuals within a population begin using different resources, they may mate more often with others using the same resource. Over time, this can reduce gene flow. For example, some insect populations become associated with different host plants. The apple maggot fly, Rhagoletis pomonella, originally used hawthorn fruit as a host. After apples were introduced to North America, some flies began using apples instead. Because these flies mate on or near their host fruit, preference for different host plants reduced gene flow between apple-associated and hawthorn-associated populations. This is an example of ecological divergence that may contribute to sympatric speciation.
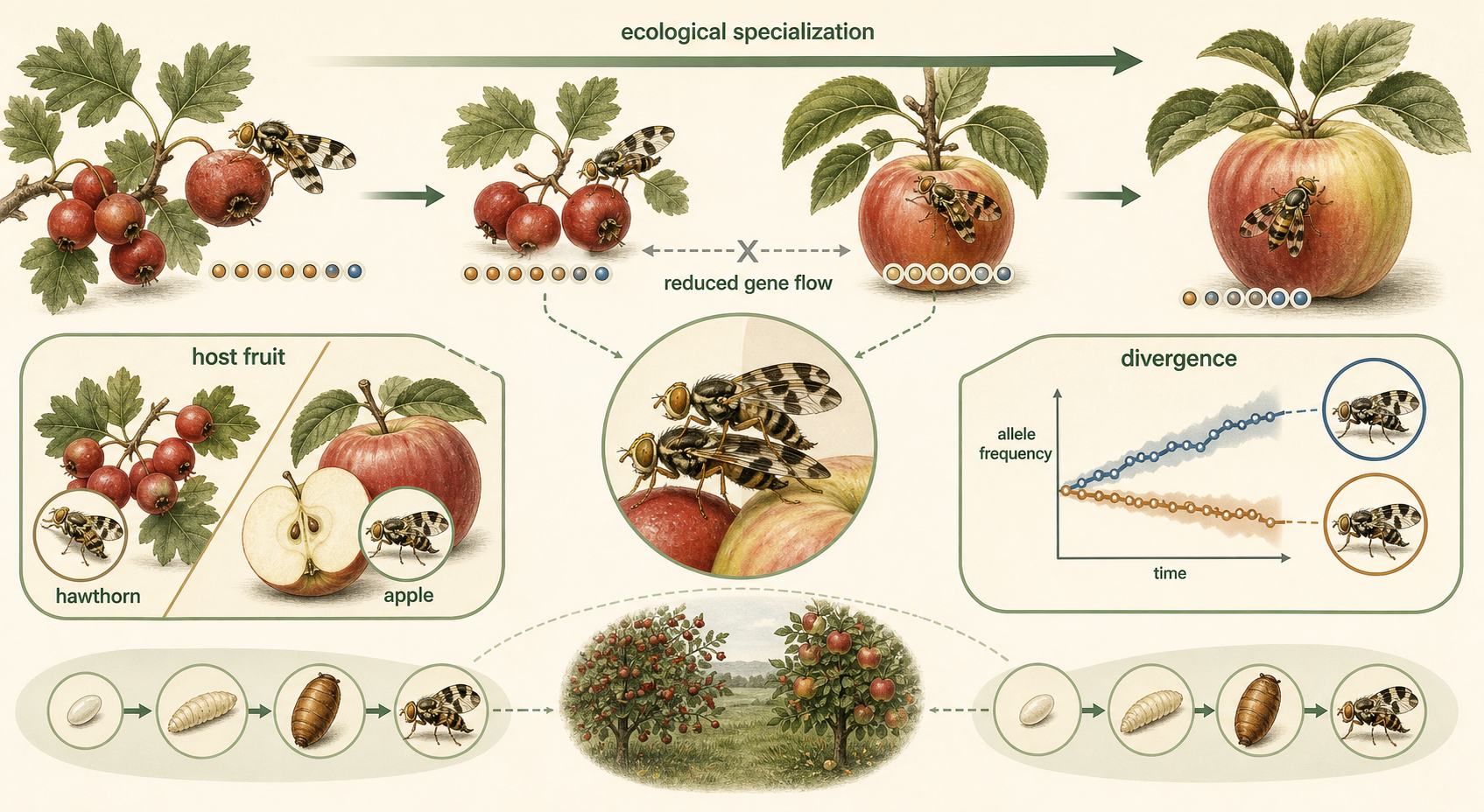
Figure 13. Sympatric Speciation by Ecological Specialization. Ecological specialization can reduce gene flow when individuals use different resources and mate with others using the same resource. Apple maggot flies illustrate how host preference can promote divergence within the same geographic area.
Sympatric Speciation: Sexual Selection
Another pathway to sympatric speciation is sexual selection. If individuals begin choosing mates based on different traits, such as color, song, size, scent, or courtship behavior, mating may become nonrandom. Over time, groups with different mating preferences may become genetically distinct, even if they live in the same place. Sexual selection can be especially important in animals that use visual signals, sounds, or elaborate courtship displays.
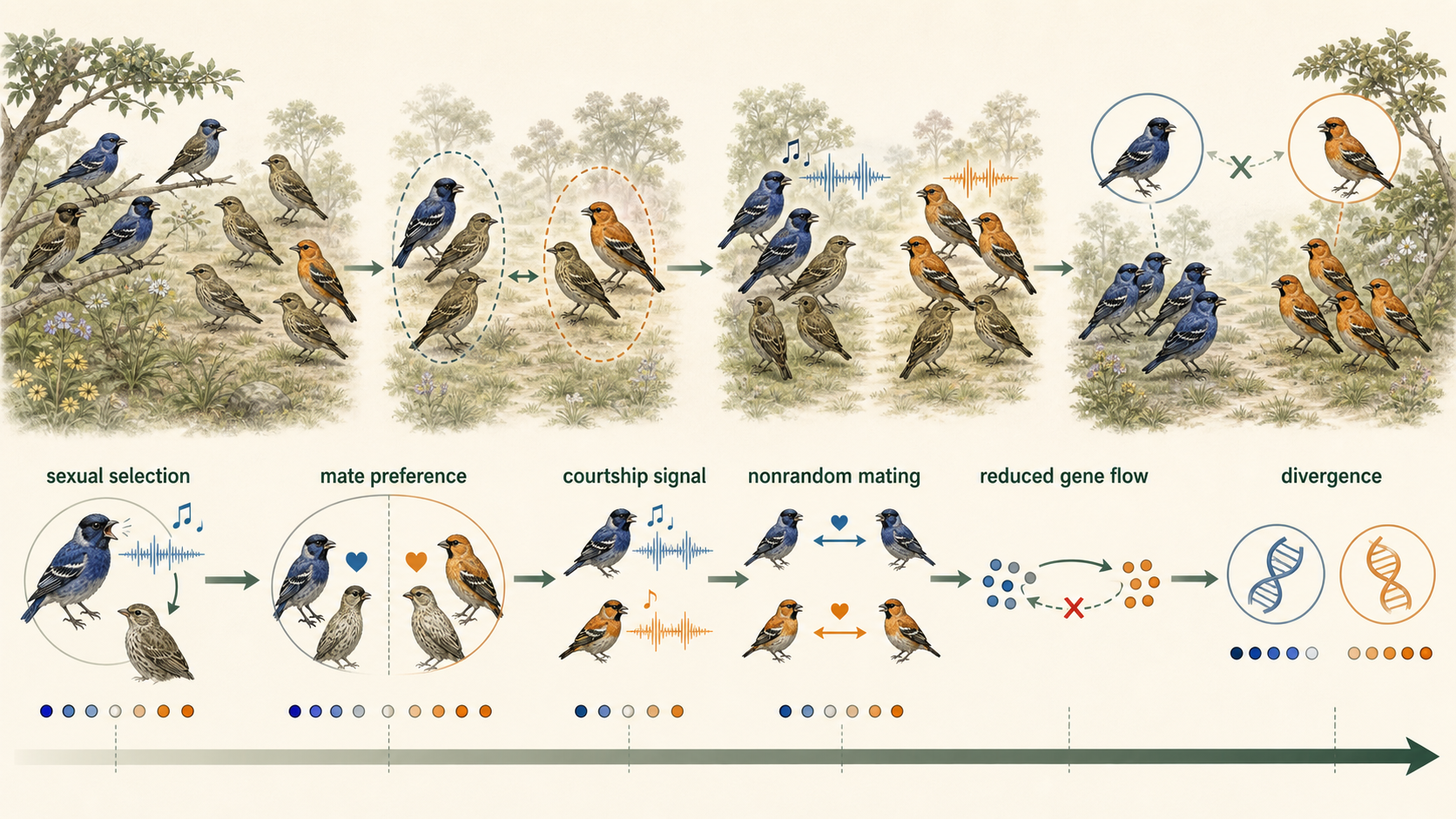
Figure 14. Sympatric Speciation by Sexual Selection. Sexual selection can promote sympatric speciation when different mating preferences reduce gene flow. Over time, groups using different signals or courtship behaviors may become genetically distinct.
Sympatric Speciation: Polyploidy
One of the fastest forms of sympatric speciation is polyploidy. Polyploidy occurs when an organism has more than two complete sets of chromosomes. A diploid organism has two sets of chromosomes, one from each parent. A tetraploid organism has four sets. Polyploidy can arise when chromosomes duplicate but the cell fails to divide properly, or when abnormal gametes with extra chromosome sets fuse during fertilization. Polyploidy is especially important in plants. A polyploid individual may be reproductively isolated from its diploid parent population almost immediately. This is because diploid and tetraploid individuals often produce offspring with uneven chromosome numbers, which can disrupt meiosis and lead to sterility. In this way, a single chromosome-level event can create a new reproductively isolated lineage in one or a few generations. Polyploidy is much more common in plants than in animals for several reasons. Plants often tolerate chromosome duplication better than animals. Many plants can self-fertilize or reproduce vegetatively, making it easier for a new polyploid individual to establish a population. Plants also hybridize relatively often, which increases the opportunity for allopolyploid speciation. Seedless watermelons provide a practical example of chromosome number affecting fertility. They are typically triploid, meaning they have three sets of chromosomes. Triploid plants are produced by crossing diploid and tetraploid watermelons. Because three chromosome sets do not pair evenly during meiosis, triploid watermelons are usually sterile and do not produce normal mature seeds. This is why seedless watermelons contain only small, undeveloped seed coats.
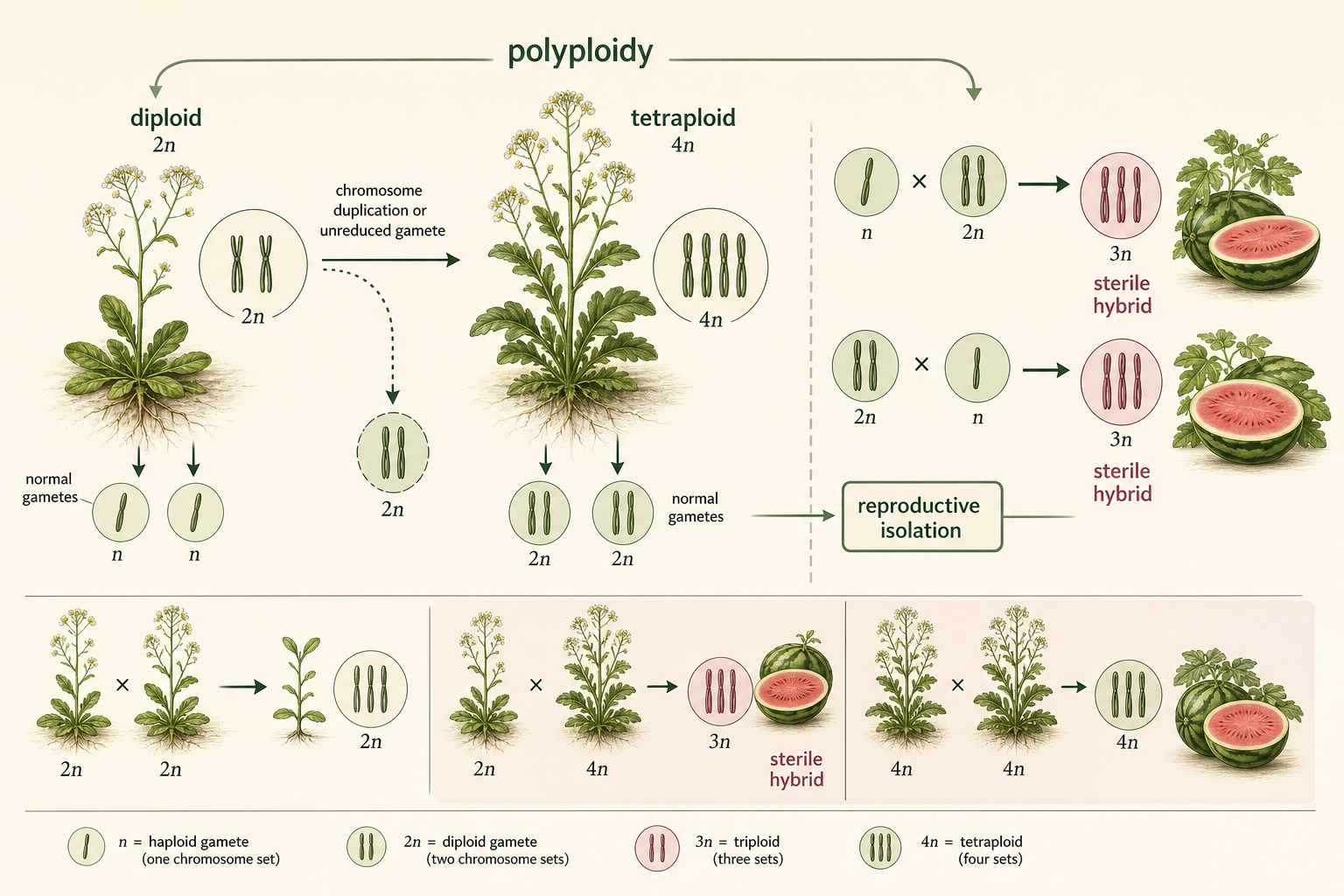
Figure 15. Sympatric Speciation by Polyploidy. Polyploidy occurs when an organism has more than two complete sets of chromosomes. In plants, chromosome duplication can rapidly create reproductive isolation from the original diploid population.
Sympatric Speciation: Polyploidy: Autopolyploidy and Allopolyploidy
There are two major types of polyploidy: autopolyploidy and allopolyploidy. Autopolyploidy occurs when chromosome sets multiply within a single species. For example, a diploid plant may produce unreduced gametes, meaning gametes that accidentally retain the full diploid chromosome number instead of becoming haploid. If two unreduced gametes fuse, the result may be a tetraploid offspring. That tetraploid may be fertile with other tetraploids but reproductively isolated from the original diploid population. Allopolyploidy occurs when two different species hybridize and chromosome duplication follows. The initial hybrid may be sterile because the chromosomes from the two parent species do not pair properly during meiosis. However, if chromosome duplication occurs, each chromosome may gain a matching partner. The result can be a fertile hybrid lineage with chromosome sets from both parent species. Many crop plants and ornamental plants have histories involving allopolyploidy.
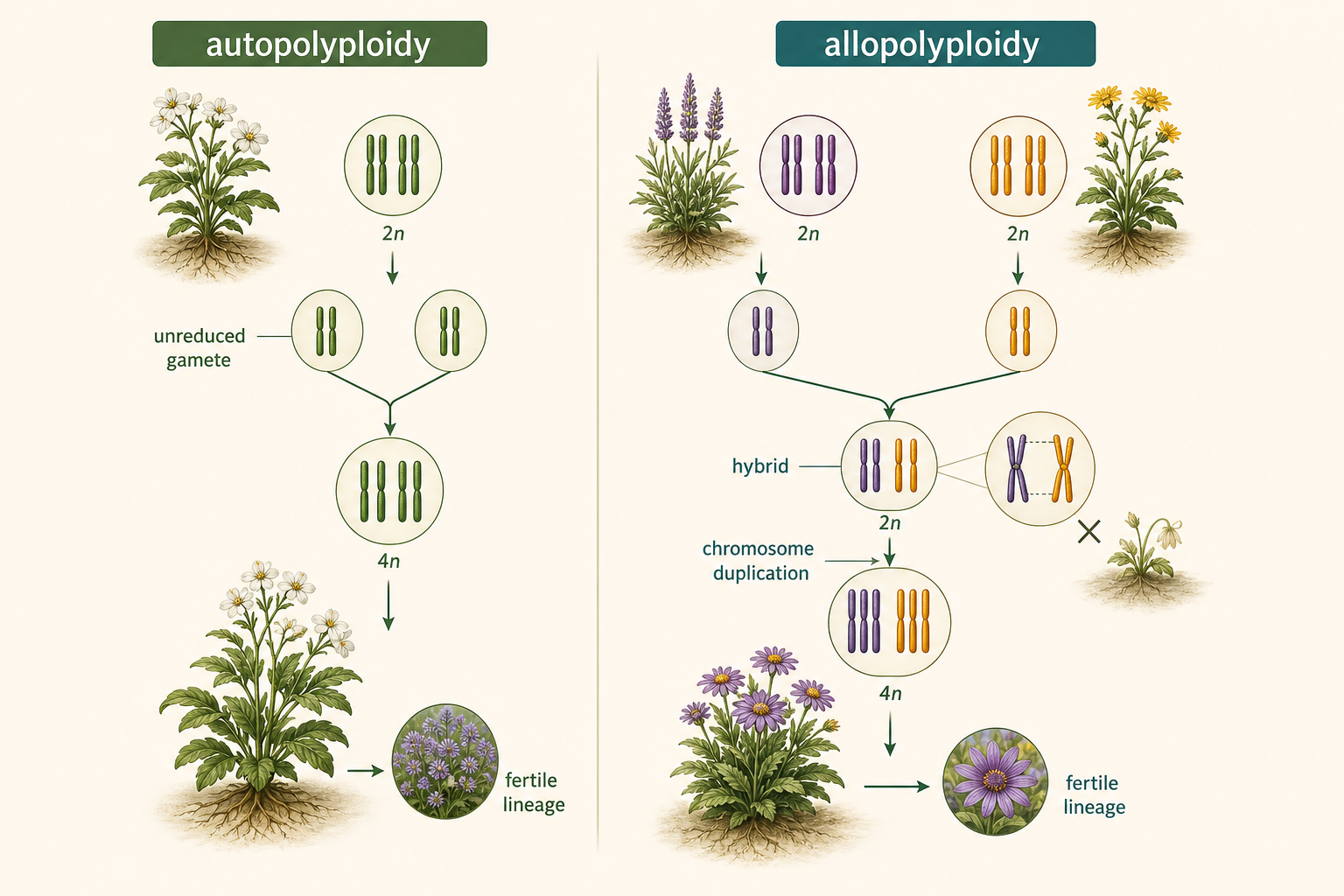
Figure 16. Autopolyploidy and Allopolyploidy. Autopolyploidy occurs when chromosome sets multiply within a single species. Allopolyploidy occurs when two species hybridize and chromosome duplication restores chromosome pairing, producing a fertile hybrid lineage.
Hybrid Zones
When separated populations or closely related species come back into contact, several outcomes are possible. If they have not diverged much, gene flow may resume and the populations may merge. If they have diverged strongly, reproductive barriers may prevent interbreeding, and the populations may remain separate species. If they can still interbreed, a hybrid zone may form. A hybrid zone is a geographic area where individuals from different species or genetically distinct populations meet and produce hybrid offspring. Hybrid zones are important because they allow scientists to study speciation in progress. They show whether reproductive barriers are weak, strong, or still evolving. Hybrid zones can have several outcomes. One outcome is fusion. If hybrids are healthy and fertile, gene flow may erase the differences between the two populations, causing them to merge back into one species. A second outcome is stability. The hybrid zone may persist for long periods if hybrids continue to be produced but do not spread far beyond the contact zone. A third outcome is reinforcement. Reinforcement occurs when hybrids have low fitness, causing natural selection to favor individuals that avoid mating with members of the other population. Over time, prezygotic barriers such as mate preference, courtship behavior, or timing of reproduction become stronger. A fourth outcome is hybrid speciation. In some cases, hybrids are not less fit. Instead, they may possess new combinations of traits that allow them to survive in a new or unusual environment. If hybrids become reproductively isolated from both parent species, they may form a new species. Hybrid speciation is especially common in plants, particularly when combined with polyploidy, but it can also occur in some animals.
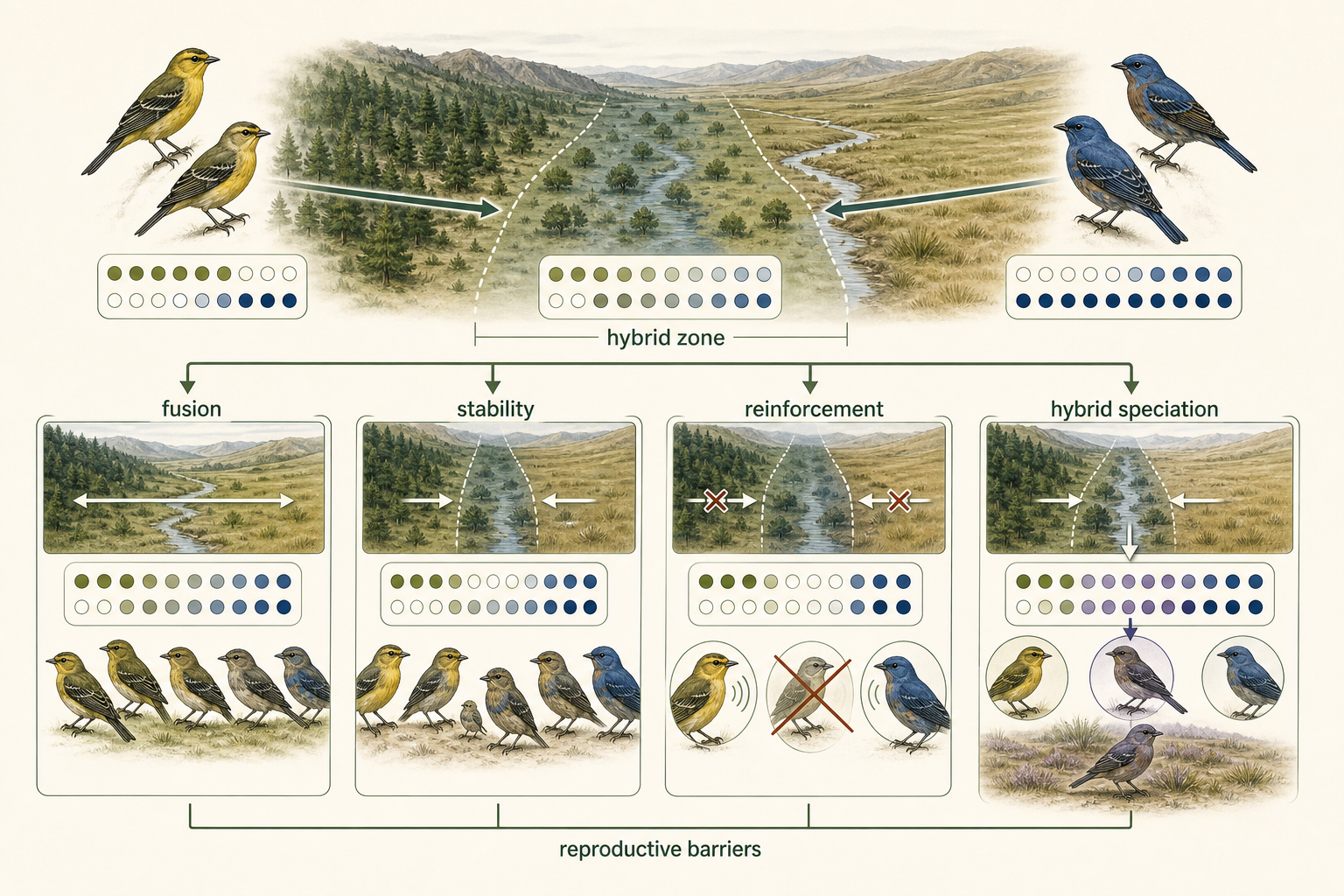
Figure 17. Hybrid Zones. A hybrid zone forms where genetically distinct populations or closely related species meet and produce hybrids. Hybrid zones may lead to fusion, stability, reinforcement, or hybrid speciation.
Speciation as a Process
Speciation is therefore not a single event with one simple cause. It is a process that unfolds as populations become genetically, ecologically, behaviorally, or reproductively distinct. Sometimes speciation is slow, taking thousands or millions of years. Sometimes it can happen rapidly, especially through polyploidy or strong ecological selection. Sometimes it stops before completion if gene flow resumes. Sometimes lineages remain partially connected through hybridization. The central theme is reproductive isolation. New species form when populations become independent evolutionary lineages. Geographic barriers, ecological specialization, genetic drift, natural selection, sexual selection, hybridization, and chromosome changes can all contribute to this process. Speciation explains how life’s diversity branches over time, producing the enormous variety of organisms found on Earth today.
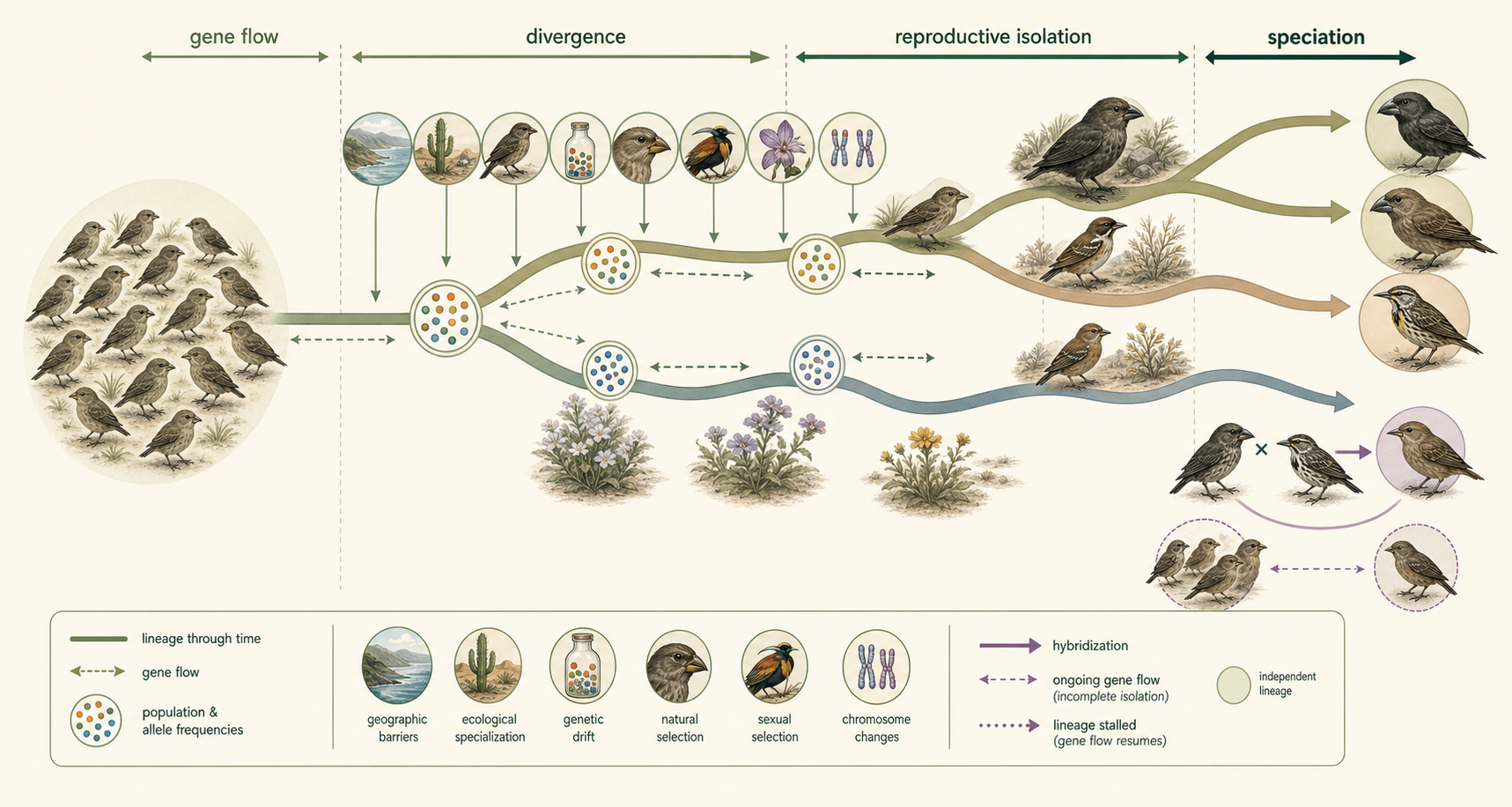
Figure 18. Speciation as a Process. Speciation is not a single event with one cause. It unfolds as populations become genetically, ecologically, behaviorally, or reproductively distinct until they become independent evolutionary lineages.