
Chapter: Eukarya
Characteristics of Eukarya
Domain Eukarya includes all organisms whose cells have a membrane-bound nucleus and other internal membrane-bound structures called organelles. This domain includes animals, fungi, plants, algae, and many single-celled organisms traditionally called protists, defined as single-celled eukaryotes. Although people often think of eukaryotes as large multicellular organisms, most eukaryotic diversity is microbial. The defining feature of eukaryotic cells is compartmentalization. In eukaryotes, DNA is housed inside the nucleus, mitochondria carry out much of cellular respiration, the endoplasmic reticulum and Golgi apparatus help process and transport molecules, lysosomes and vacuoles help digest or store materials, and the cytoskeleton supports cell shape and movement. These internal compartments allow different cellular processes to occur in different places at the same time. This made eukaryotic cells larger, more structurally complex, and more flexible than prokaryotic cells. Eukaryotes also usually have linear chromosomes, complex gene regulation, cytoskeletal movement, and sexual reproduction involving meiosis. Together, these traits helped make eukaryotes one of the major branches of life on Earth.
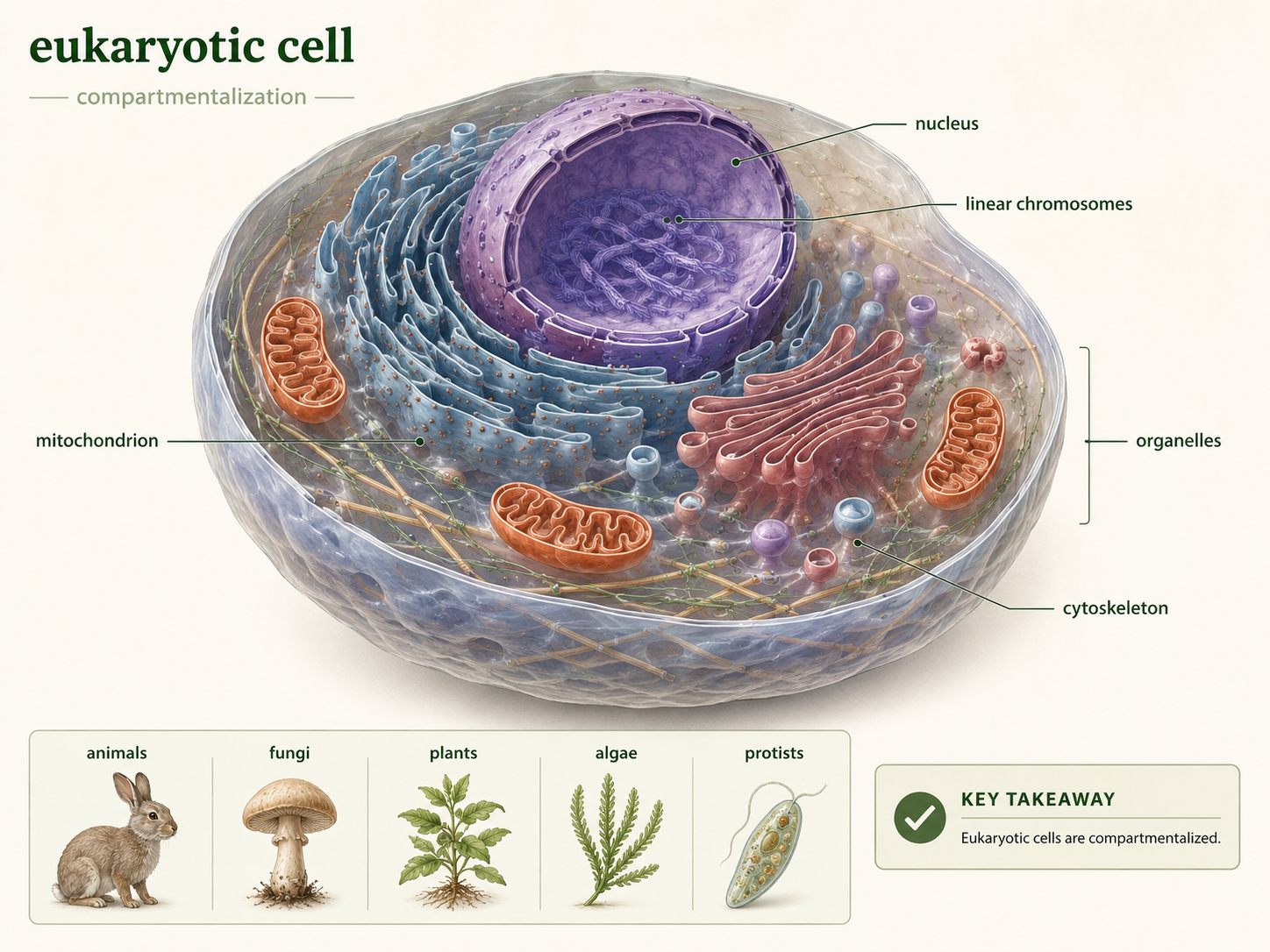
Figure 1. Characteristics of Eukarya. Eukaryotes are organisms with cells that contain a nucleus, linear chromosomes, membrane-bound organelles, a cytoskeleton, and complex gene regulation. The group includes unicellular protists as well as multicellular fungi, animals, plants, and algae.
Eukarya and the Tree of Life
Domain Eukarya is one of the major divisions of life, along with Bacteria and Archaea. Traditional biology often presents life as three domains: Bacteria, Archaea, and Eukarya. This model is still useful for introductory biology because it highlights the major differences between cells with nuclei and cells without nuclei. However, modern evolutionary evidence suggests that eukaryotes are especially closely related to Archaea. In other words, eukaryotes did not simply appear as a completely separate kind of cell. Domain Eukarya likely evolved from an archaeal ancestor. This origin helps explain why eukaryotic cells are biologically mixed. Their information-processing systems, such as transcription and translation, resemble those of archaea in several ways. Their mitochondria, however, originated from bacteria through endosymbiosis (more on that later). Eukaryotic cells therefore carry evidence of more than one evolutionary source. They are not just “larger prokaryotes.” They are cells shaped by membrane evolution, genetic innovation, cytoskeletal complexity, and ancient symbiotic relationships.
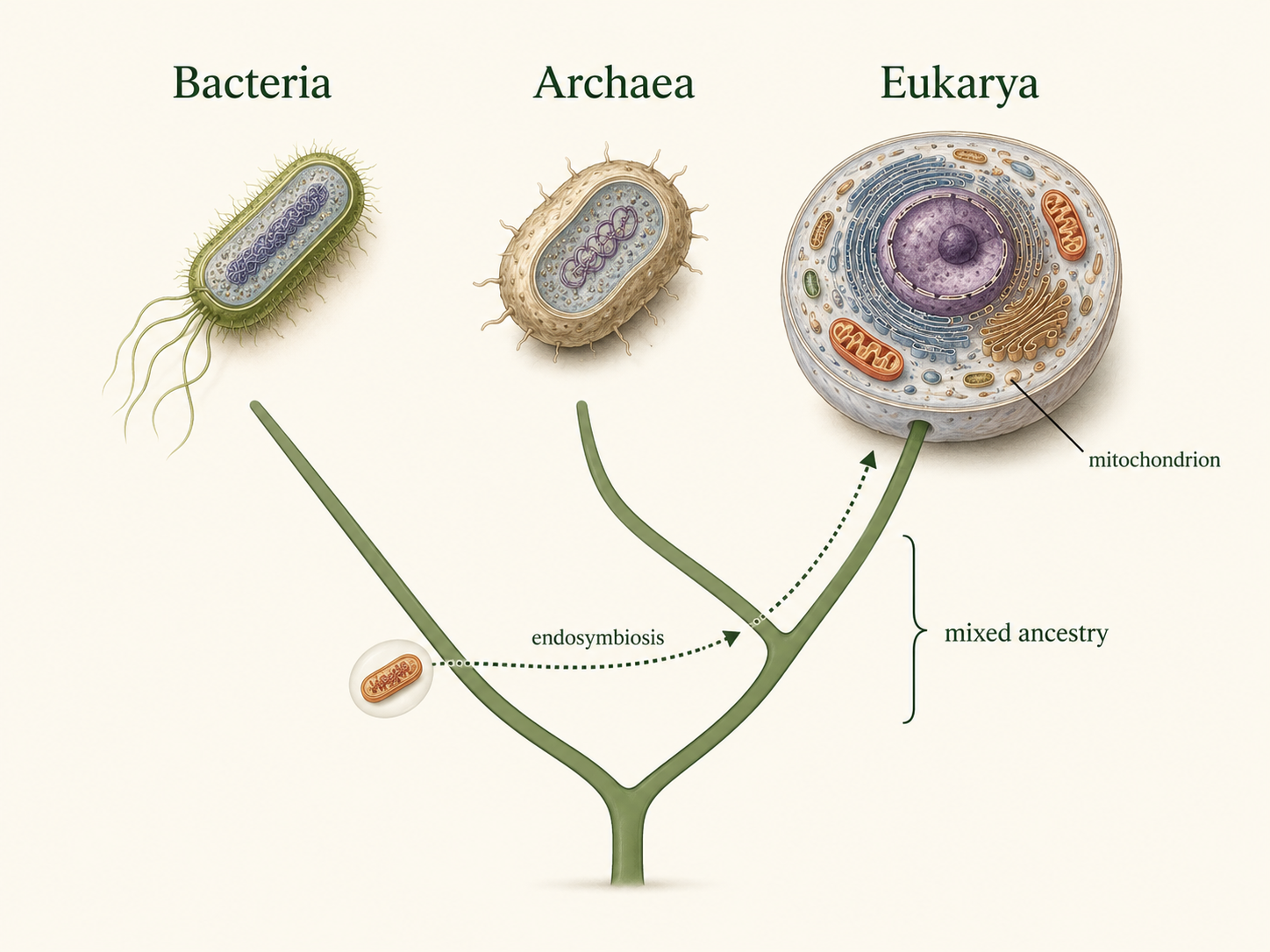
Figure 2. Eukarya and the tree of life. Eukaryotes are closely related to Archaea but also contain bacterial-derived organelles, especially mitochondria. This mixed ancestry reflects the deep evolutionary events that produced complex eukaryotic cells.
The Origin of the Nucleus
Domain Eukarya
The nucleus is the structure that gives eukaryotes their name. The word eukaryote means “true nucleus.” The nucleus stores the cell’s DNA inside a double membrane called the nuclear envelope. This separates DNA from the cytoplasm, where ribosomes translate messenger RNA into proteins. In prokaryotic cells, transcription and translation can happen at the same time because there is no nucleus separating DNA from ribosomes. In eukaryotic cells, transcription occurs in the nucleus, RNA is processed, and then mature messenger RNA exits through nuclear pores to be translated in the cytoplasm. One major hypothesis proposes that the nucleus originated from infoldings of the cell membrane. In an early archaeal-like cell, sections of the cell membrane may have folded inward, surrounded the DNA, and eventually formed an internal membrane system. Over time, these membranes became the nuclear envelope. The advantage of this arrangement was not simply storage. The nucleus allowed more complex gene regulation. Because RNA could be modified before translation, eukaryotic cells gained new ways to control when and how genes were expressed. This separation of transcription and translation became one of the most important innovations in eukaryotic evolution.
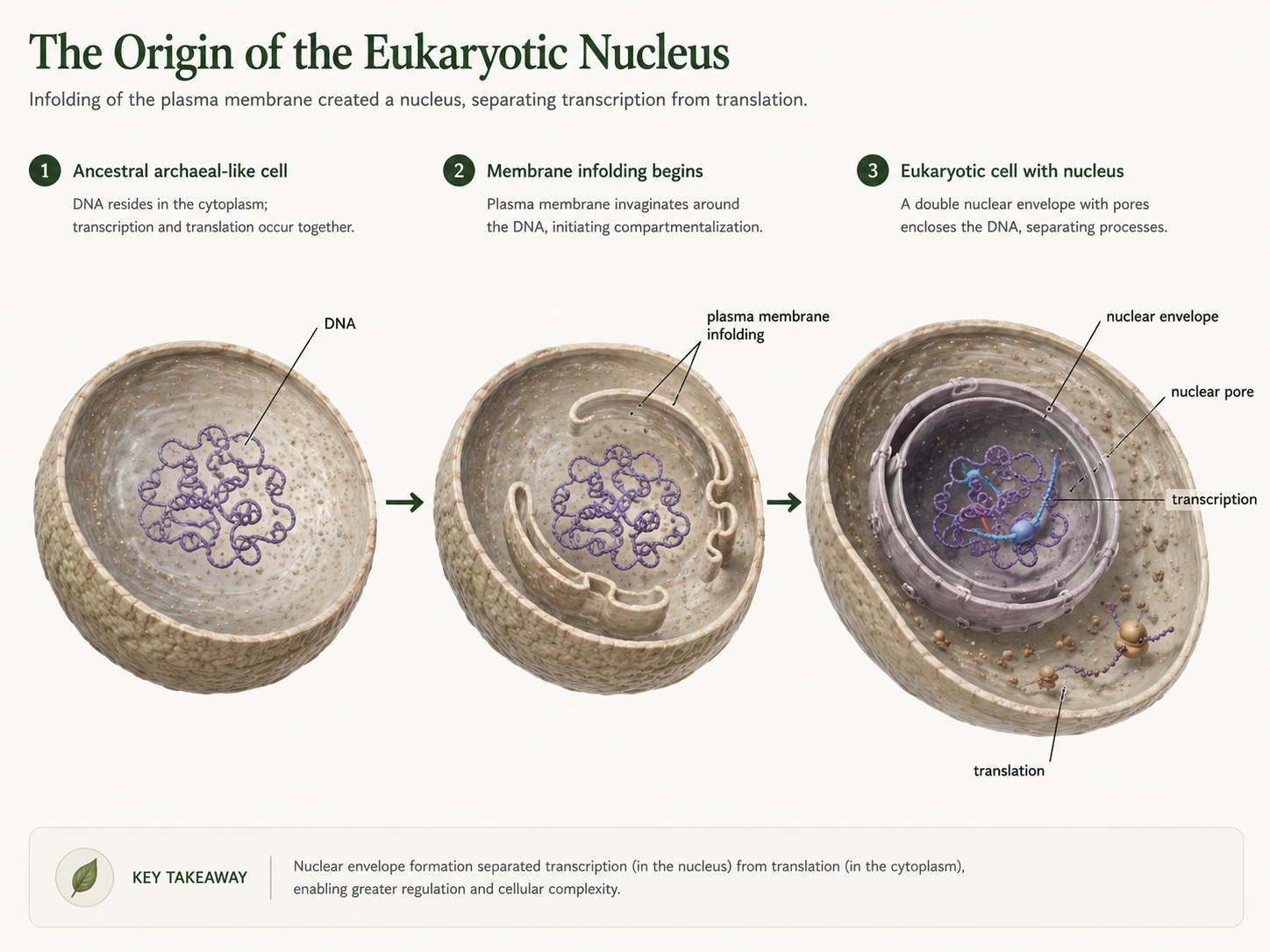
Figure 3. Origin of the nucleus. One hypothesis proposes that the nuclear envelope evolved from infoldings of the plasma membrane that gradually surrounded the cell’s DNA. This created a separate compartment for gene storage and regulation, allowing transcription and translation to be separated.
The Endomembrane System
Domain Eukarya
The origin of the nucleus actually includes a larger internal membrane system known as the endomembrane system. This system includes the nucleus, endoplasmic reticulum, Golgi apparatus, lysosomes, vacuoles, and vesicles. The endomembrane system likely evolved through repeated folding, specialization, and compartmentalization of internal membranes. Once internal membranes existed, different regions could specialize for different tasks. The rough endoplasmic reticulum became associated with ribosomes and protein production. The smooth endoplasmic reticulum became important in lipid synthesis and detoxification. The Golgi apparatus modified and sorted molecules. Lysosomes and vacuoles became involved in digestion, recycling, and storage. This internal organization allowed eukaryotic cells to become larger and more complex without losing control over their internal chemistry.
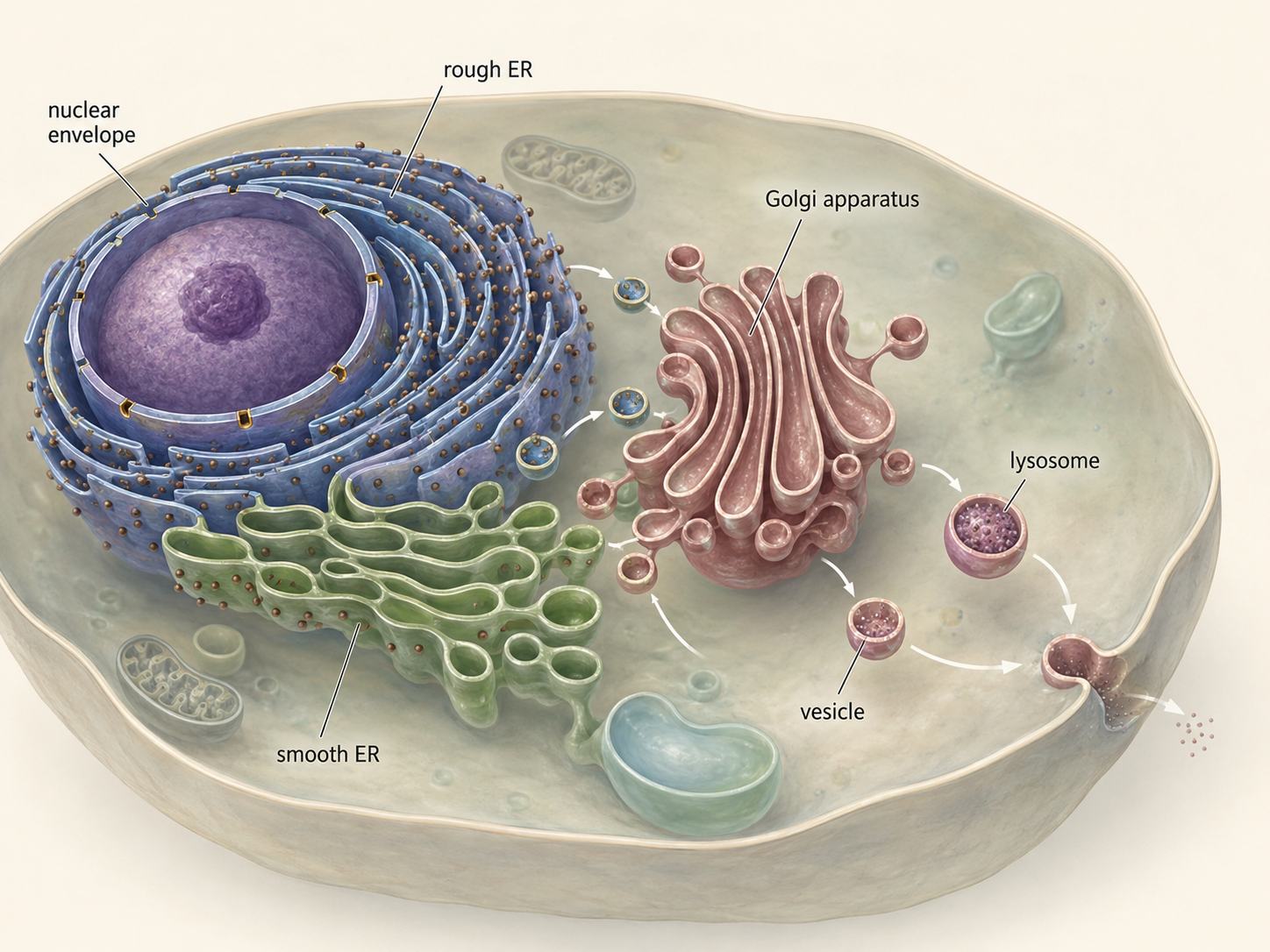
Figure 4. The endomembrane system. The endomembrane system includes the nuclear envelope, endoplasmic reticulum, Golgi apparatus, vesicles, lysosomes, vacuoles, and plasma membrane. These compartments allow eukaryotic cells to process, transport, digest, and organize materials more efficiently.
The Mitochondrion: Endosymbiosis and Energy
Domain Eukarya
Mitochondria are organelles that help eukaryotic cells harvest energy from organic molecules. Their origin is one of the strongest examples of endosymbiosis, an evolutionary process in which one organism lives inside another. According to the mitochondrial endosymbiotic hypothesis, an early eukaryotic cell engulfed a bacterium but did not digest it. Instead, the bacterium survived inside the host cell. Over time, the host cell provided protection and nutrients, while the internal bacterium provided a more efficient way to extract energy from food. This internal bacterium eventually evolved into the mitochondrion. Several lines of evidence support this hypothesis. First, mitochondria have their own DNA, and mitochondrial genes are most closely related to bacterial genes. Second, mitochondria also divide in a way that resembles bacterial cell division, binary fission. Third, they have a double membrane, consistent with an engulfment event, and their ribosomes resemble bacterial ribosomes more than eukaryotic cytoplasmic ribosomes. Modern mitochondria are not independent bacteria because many genes have moved to the nucleus, and mitochondria depend on the host cell for many proteins. Still, their structure and genetics preserve evidence of their bacterial origin. The evolution of mitochondria was a major turning point in the history of life. The obvious advantage of cells with mitochondria is that they can produce much more ATP from food than cells relying only on fermentation. This increased energy availability may have helped support larger cell size, active transport, internal membranes, cytoskeletal movement, and eventually multicellularity.
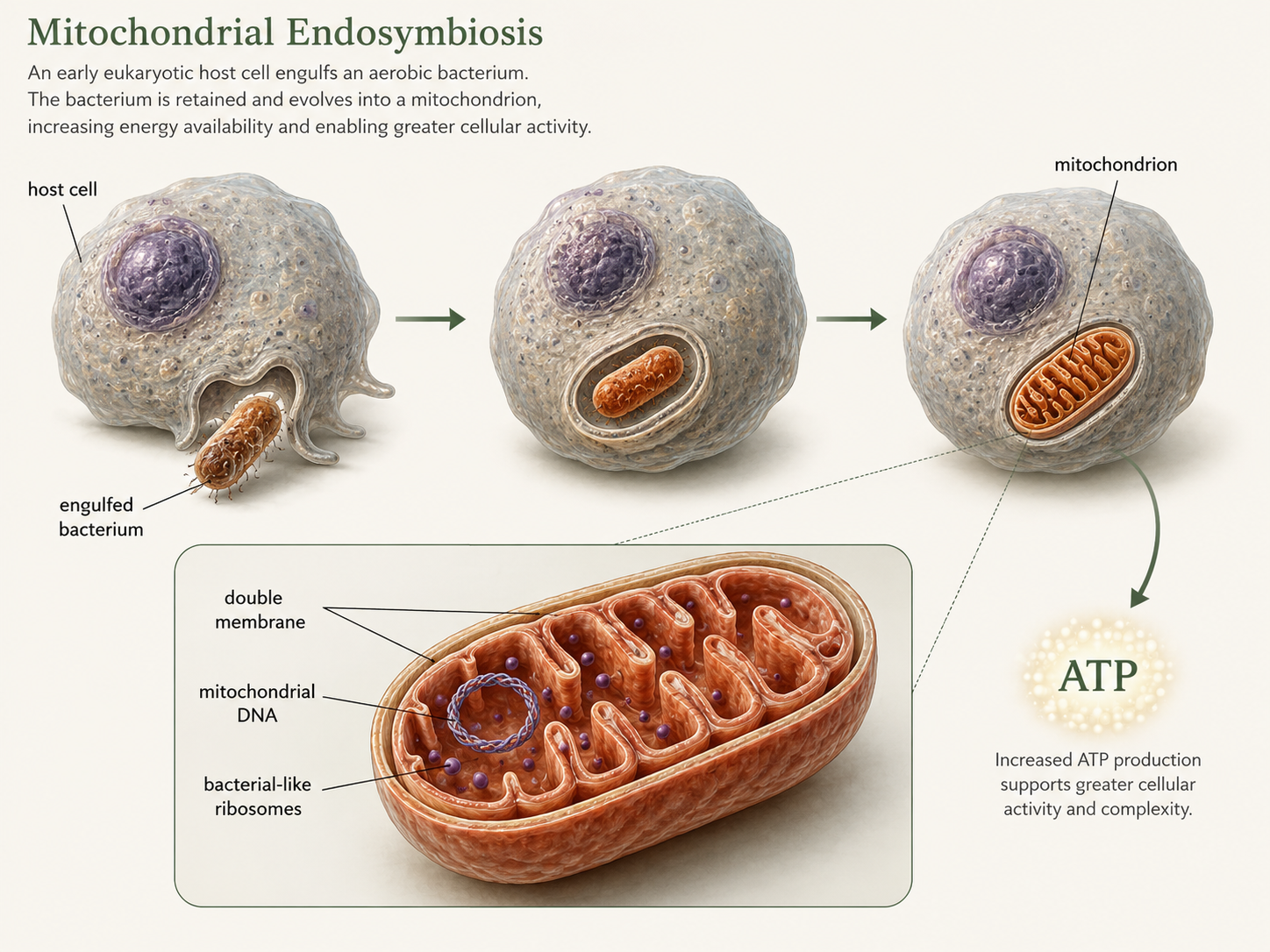
Figure 5. Mitochondrial endosymbiosis. Mitochondria evolved when an early eukaryotic host cell engulfed a bacterium that was not digested. The bacterial symbiont eventually became an organelle that helped the host cell produce ATP more efficiently. Mitochondrial DNA, bacterial-like ribosomes, division by fission, and a double membrane support this hypothesis.
The Origin of Chloroplasts
Domain Eukarya: Archaeplastida
Chloroplasts are photosynthetic organelles that allow many eukaryotes to capture light energy and use it to build sugars. Like mitochondria, chloroplasts evolved through endosymbiosis. In this case, the engulfed organism was a cyanobacterium, a photosynthetic bacterium capable of oxygen-producing photosynthesis. An early eukaryotic cell engulfed a cyanobacterium but did not digest it. The cyanobacterium produced sugars through photosynthesis, while the host cell provided protection and access to resources. Over time, this cyanobacterium evolved into the chloroplast. This event is called primary endosymbiosis because a eukaryotic cell directly engulfed a cyanobacterium. The descendants of this event include the Archaeplastida, a group that includes glaucophyte algae, red algae, green algae, and land plants. These organisms have plastids that trace back to a cyanobacterial ancestor. Like mitochondria evidence exists of this primary endosymbiotic event: 1) chloroplasts have their own DNA, 2) bacterial-like ribosomes, 3) double membranes, and 4) chloroplast genes show a close evolutionary relationship to cyanobacteria. The origin of chloroplasts changed Earth’s ecosystems. Photosynthetic eukaryotes became major producers in aquatic environments, forming the base of many food webs. Later, one branch of green algae gave rise to land plants, which transformed terrestrial ecosystems by producing oxygen, storing carbon, forming soils, and supporting animal life on land.
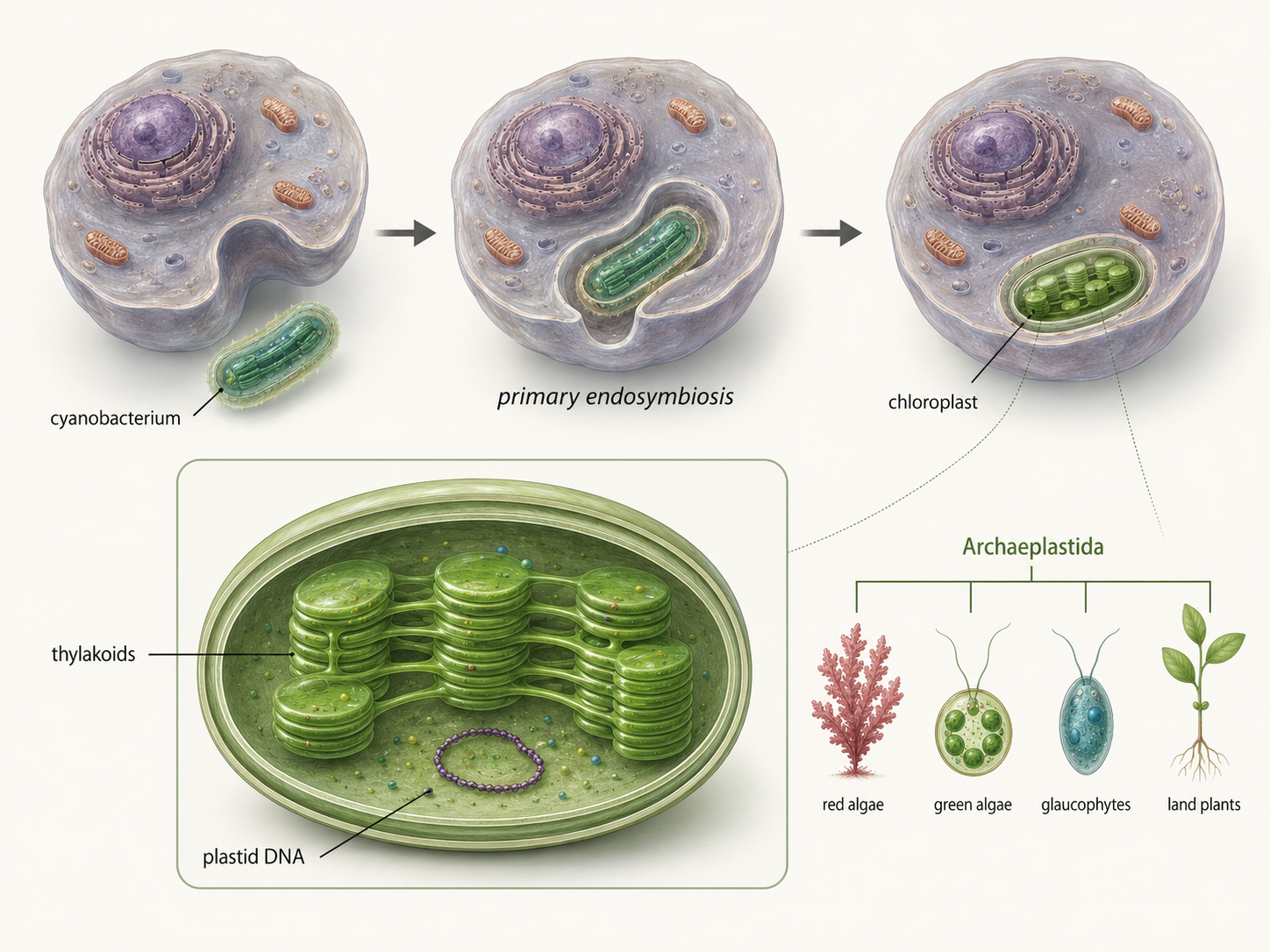
Figure 6. Primary endosymbiosis of the chloroplast. Chloroplasts evolved when an early eukaryotic cell engulfed a photosynthetic cyanobacterium. This event gave rise to the plastids of Archaeplastida, including red algae, green algae, glaucophytes, and land plants.
Secondary Endosymbiosis: Cells Inside Cells
Domain Eukarya
The history of chloroplasts did not end with primary endosymbiosis. In several lineages, one eukaryote engulfed another eukaryote that already contained a chloroplast. This process is called secondary endosymbiosis. It can be thought of as a cell engulfing a cell that had already engulfed a cell. These events helped spread photosynthesis across distantly related eukaryotic groups. Secondary endosymbiosis explains why some photosynthetic eukaryotes have plastids surrounded by more than two membranes. The innermost membranes come from the original cyanobacterium. Additional membranes come from the engulfed alga and from the host cell that captured it. In some lineages, tiny remnants of the engulfed algal nucleus, called nucleomorphs, remain between plastid membranes. These structures provide strong evidence that the plastid passed through a eukaryotic algal stage before becoming part of another eukaryotic cell. Secondary endosymbiosis occurred more than once. Euglenids acquired plastids from green algae. Many stramenopiles, such as diatoms and brown algae, acquired plastids ultimately derived from red algae. Some alveolates, including dinoflagellates and apicomplexans, also carry evidence of red algal plastid ancestry, although some have lost photosynthesis. These repeated events make eukaryotic evolution difficult to draw as a simple tree. In many cases, it is better imagined as a tree with branches connected by ancient cellular mergers
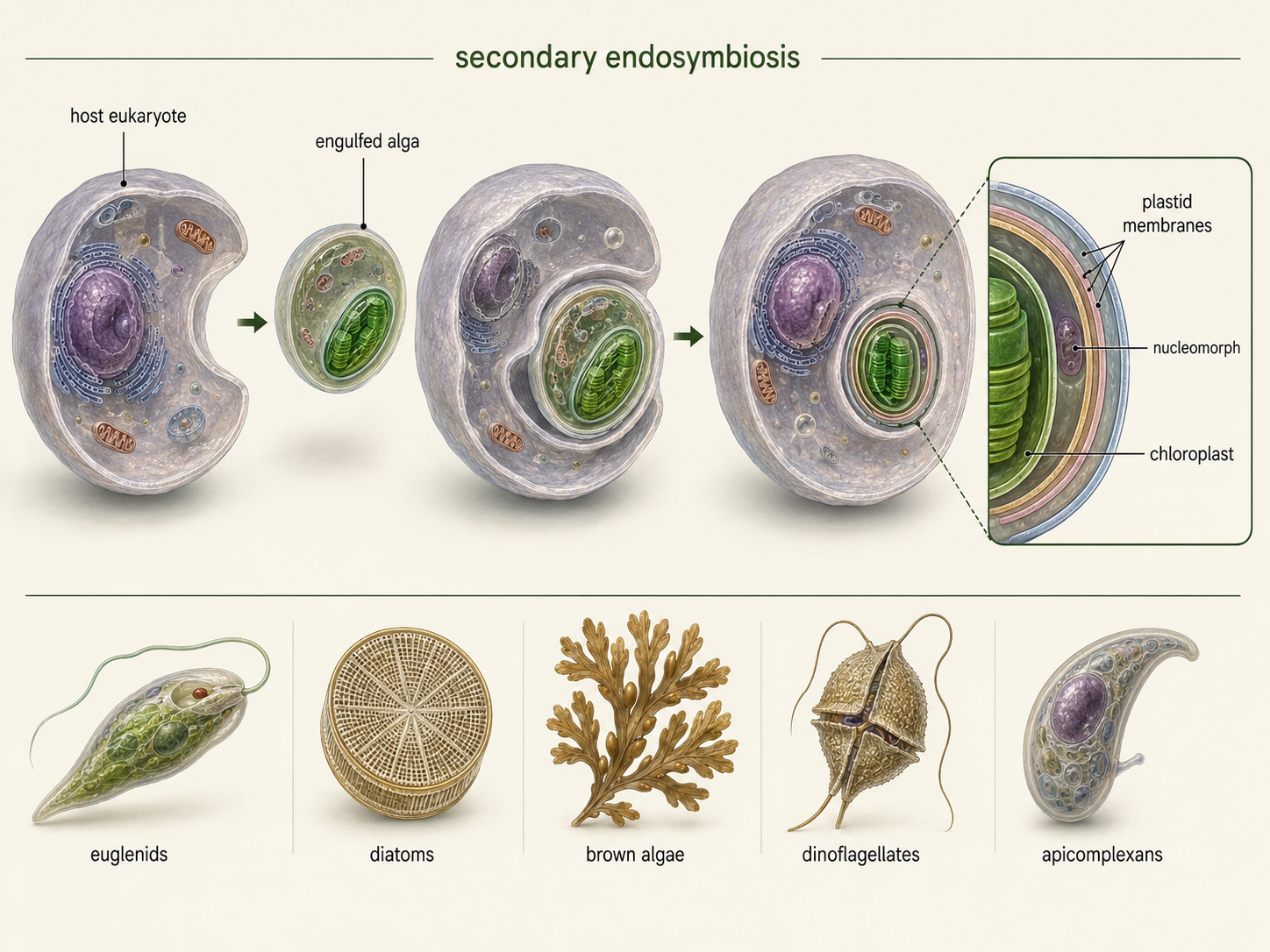
Figure 7. Secondary endosymbiosis. Secondary endosymbiosis occurs when a eukaryote engulfs another eukaryote that already contains a chloroplast. This process spread photosynthesis into several unrelated eukaryotic lineages and helps explain why some plastids have three or four surrounding membranes.
Understanding Eukaryotic Classification
Domain Eukarya
Classifying eukaryotes is difficult because much of eukaryotic diversity is microbial, ancient, and morphologically variable. Animals, fungi, and plants are familiar because they are large and multicellular, but they represent only a small part of eukaryotic diversity. Many eukaryotes are single-celled organisms that were historically grouped together as protists. The term protist is still useful in teaching, but it does not describe one natural evolutionary group. A protist is generally any eukaryote that is not an animal, land plant, or fungus.
Older textbooks often divided Eukarya into six supergroups: Excavata, Amoebozoa, Opisthokonta, Rhizaria, Chromoalveolata, and Archaeplastida. This model was useful because it gave students a manageable way to organize eukaryotic diversity. However, some of these groupings have changed as DNA sequencing and phylogenomics have improved. Chromoalveolata, in particular, is no longer treated as a strong single clade in many modern classifications. Many organisms once placed there are now taught within SAR, a major grouping that includes Stramenopiles, Alveolates, and Rhizaria. Excavata is also treated cautiously because its members may not form one natural clade. For an introductory biology course, the most important point is not to memorize one perfect eukaryotic tree. The deeper relationships among eukaryotes are still being revised. The stronger goal is to understand the major lineages, the traits that help identify them, and the evolutionary events that produced their diversity.
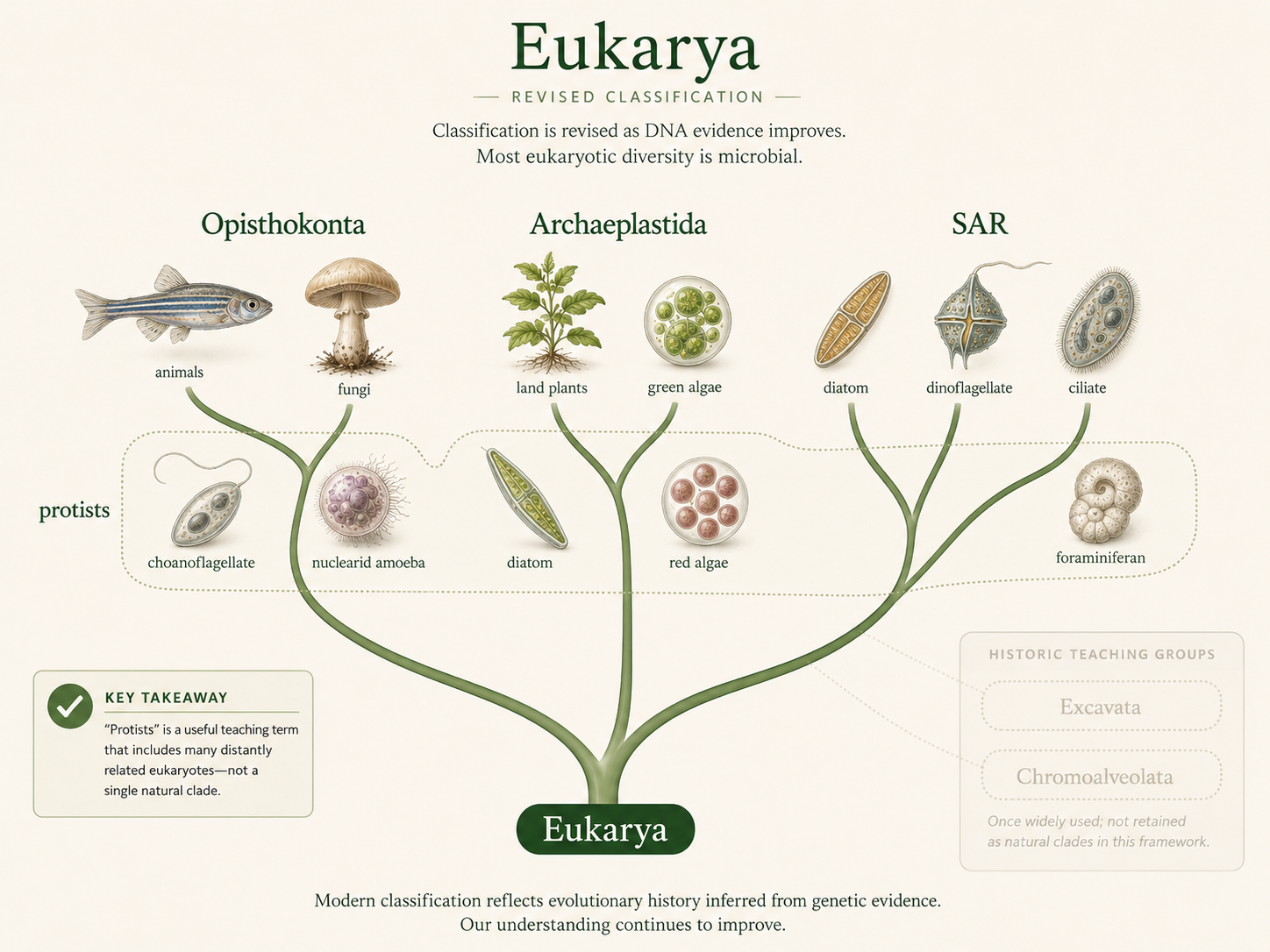
Figure 8. Classification of Eukarya. Eukaryotic classification is actively revised as new genetic and genomic evidence becomes available. Terms such as protist, Excavata, and Chromoalveolata can be useful historically or pedagogically, but they should be treated carefully because they do not always represent stable modern clades.
Excavata and Excavata-Like Lineages
Domain Eukarya: Excavata-grade lineages
Excavata has traditionally included several groups of flagellated protists, including diplomonads, parabasalids, and euglenozoans. These organisms are often discussed together because many have distinctive flagella, unusual mitochondria or mitochondrion-derived organelles, and feeding grooves or cytoskeletal supports. However, modern evidence suggests that Excavata may not be a single natural clade. For teaching purposes, it is useful to treat these organisms as excavate-like lineages rather than as one settled evolutionary group. Many excavate-like organisms are unicellular and live in aquatic, moist, or host-associated environments. Some are free-living predators of bacteria and other protists. Others are parasites. Some are photosynthetic because they acquired chloroplasts through secondary endosymbiosis. This combination of traits makes excavate-like lineages important for understanding how eukaryotic cells diversify through changes in movement, metabolism, feeding, and symbiosis.
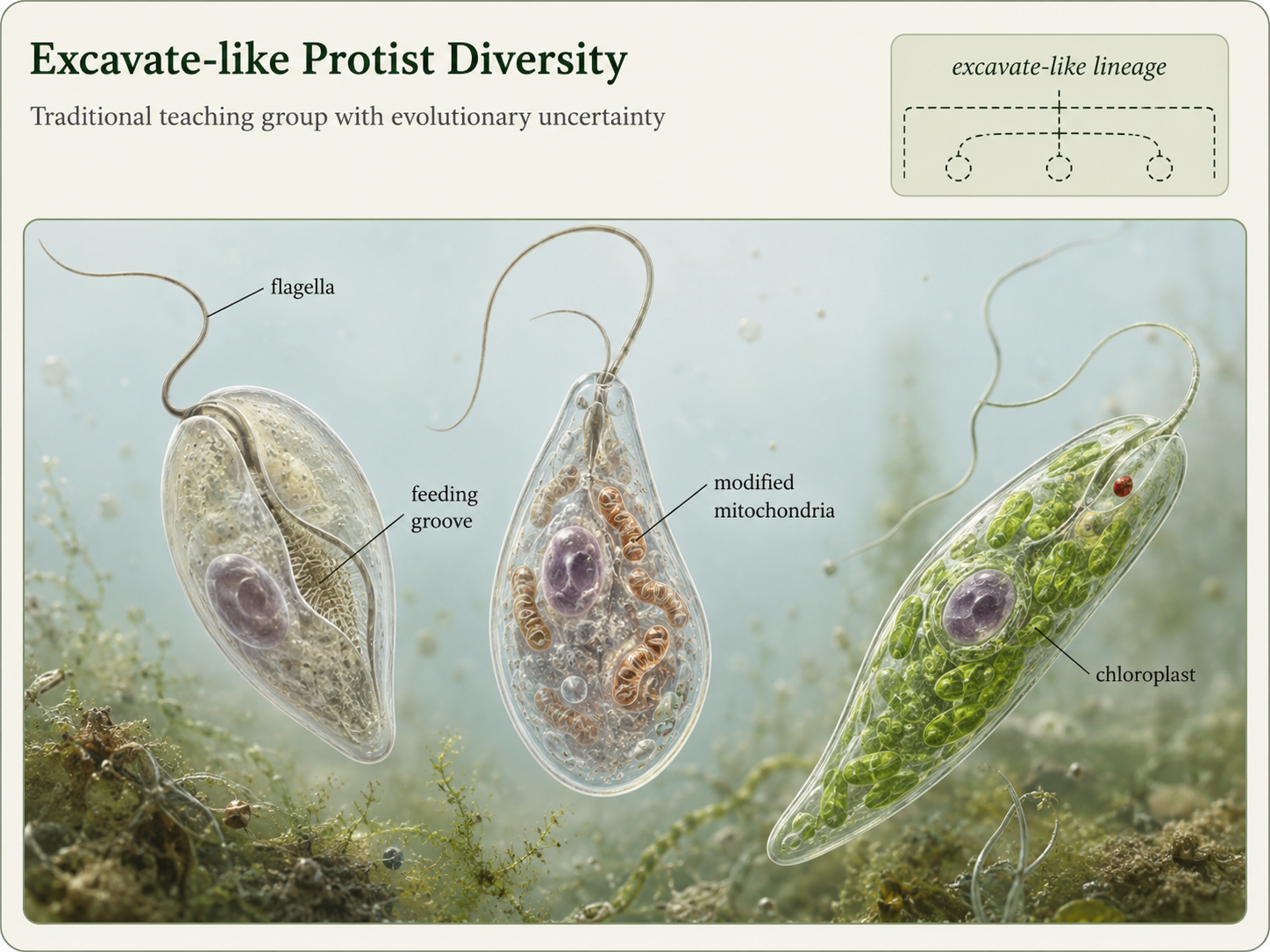
Figure 9. Excavata-like lineages. Excavata has traditionally included several flagellated protist groups, but its status as a single natural clade is uncertain. These organisms remain useful teaching examples because they show diverse forms of movement, feeding, mitochondrial modification, and secondary endosymbiosis.
Metamonads: Modified Mitochondria and Anaerobic Life
Domain Eukarya: Excavata-grade lineages: Metamonada
Metamonads include diplomonads and parabasalids, groups of unicellular flagellated protists that often live in low-oxygen environments. Many metamonads have highly modified mitochondria rather than typical energy-producing mitochondria. In diplomonads, these organelles are called mitosomes. In parabasalids, they are called hydrogenosomes. These organelles do not function like typical mitochondria in aerobic respiration, but they are still evolutionary descendants of mitochondria. Giardia is one of the best-known diplomonads. It is an intestinal parasite that can cause diarrheal disease in humans and other animals. Its simplified cellular structure once led some scientists to think it represented a very primitive eukaryote that evolved before mitochondria. Modern evidence instead supports a different explanation: Giardia and its relatives descended from ancestors that had mitochondria, but those mitochondria became highly reduced as these organisms adapted to low-oxygen environments.
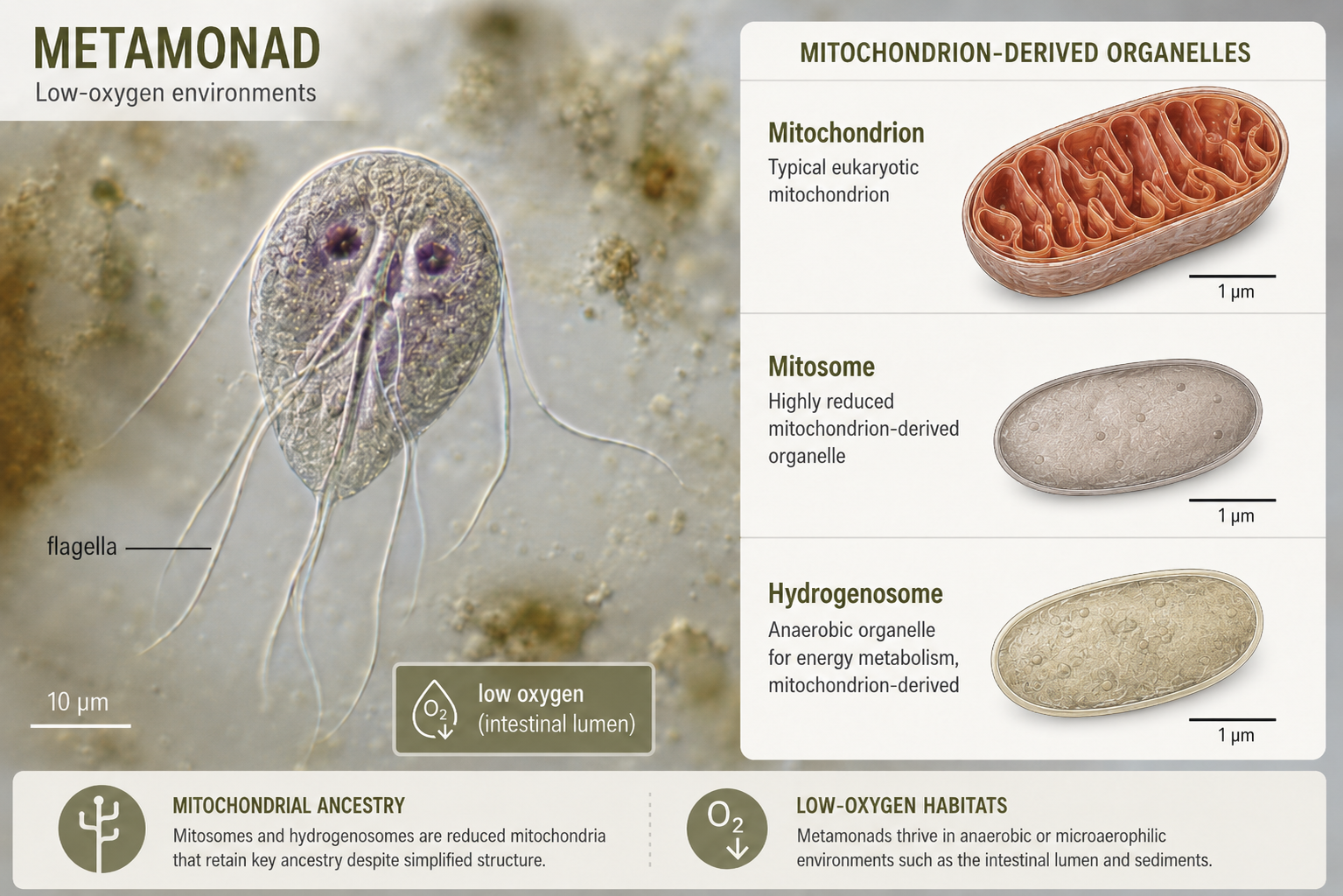
Figure 10. Metamonads. Metamonads are flagellated protists that often live in low-oxygen environments and contain modified mitochondria, such as mitosomes or hydrogenosomes. Giardia is a familiar diplomonad parasite whose simplified cell structure reflects evolutionary reduction, not the absence of mitochondrial ancestry.
Euglenozoa: Predators, Parasites, and Mixotrophs
Domain Eukarya: Excavata-grade lineages: Euglenozoa
Euglenozoa is a diverse group of flagellated protists that includes kinetoplastids and euglenids. Kinetoplastids are named for the kinetoplast, a dense region of mitochondrial DNA located within a large mitochondrion near the base of the flagellum. Some kinetoplastids are free-living predators that consume bacteria and other protists. Others are parasites, including species that cause serious diseases in humans, animals, and plants. Euglenids are especially useful for teaching because many are mixotrophs, meaning they can use more than one nutritional strategy. Some euglenids photosynthesize when light is available because their ancestors acquired chloroplasts through secondary endosymbiosis with a green alga. However, many can also feed on other organisms or absorb organic molecules. Some euglenids have an eyespot that helps them detect light and orient themselves for photosynthesis. Their flexible nutrition shows that the categories “plant-like” and “animal-like” do not always work well for unicellular eukaryotes.
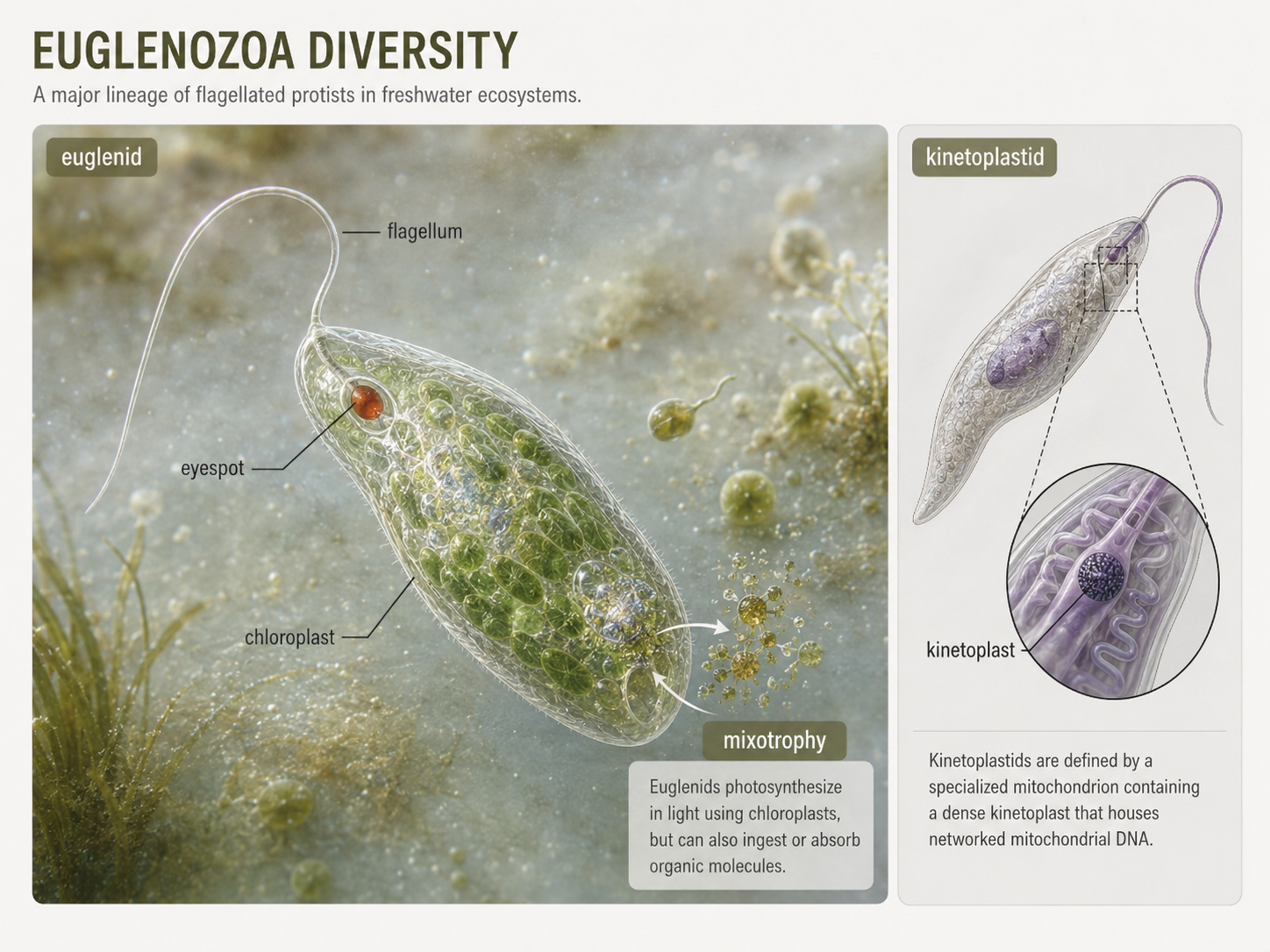
Figure 11. Euglenozoa. Euglenozoans include kinetoplastids with dense mitochondrial DNA regions and euglenids that may be photosynthetic, heterotrophic, or mixotrophic. Euglenids show why many protists do not fit neatly into plant-like or animal-like categories.
Amoebozoa: Movement by Pseudopodia
Domain Eukarya: Amoebozoa
Amoebozoans are eukaryotes that often move and feed using blunt extensions of the cell called lobose pseudopodia. A pseudopodium is a temporary extension of the cytoplasm supported by the cytoskeleton. Amoeboid cells extend a pseudopodium forward, anchor it, and then pull the rest of the cell in that direction. This movement allows many amoebozoans to crawl across surfaces or through sediment. Amoebozoans are typically heterotrophs. Many feed by phagocytosis, a process in which the cell surrounds and engulfs food particles, bacteria, or other protists. Once engulfed, the food is digested inside a vacuole. This feeding style makes amoebozoans important microbial predators in freshwater, marine, soil, and moist terrestrial ecosystems. By consuming bacteria and other microbes, they help regulate microbial communities and recycle nutrients.

Figure 12. Amoebozoa. Amoebozoans often move and feed using blunt lobose pseudopodia. These extensions allow the cell to crawl, surround prey, and engulf food through phagocytosis.
Gymnamoebas: Classic Amoebas
Domain Eukarya: Amoebozoa: Gymnamoebas
Gymnamoebas are the classic free-living amoebas found in freshwater, marine sediments, soils, and moist terrestrial habitats. They lack a rigid external shell and usually move by extending lobose pseudopodia. Many are active predators of bacteria, algae, and other protists. Their flexible shape allows them to squeeze through small spaces, surround food, and respond quickly to their environment. Although gymnamoebas are unicellular, they are not simple in a functional sense. Their movement requires coordinated cytoskeletal rearrangement, and their feeding requires membrane remodeling, prey capture, digestion, and waste removal. They are excellent examples of how a single eukaryotic cell can behave as a complete organism.

Figure 13. Gymnamoebas. Gymnamoebas are free-living amoebas that move with lobose pseudopodia and feed by phagocytosis. They show how a single eukaryotic cell can move, hunt, digest, and respond to its environment.
Slime Molds: Unicellular Life and Cooperative Bodies
Domain Eukarya: Amoebozoa: Mycetozoa
Slime molds are amoebozoans that challenge simple definitions of unicellularity and multicellularity. Many slime molds spend part of their life cycle as individual cells. When food is abundant, these cells may live independently, moving through soil or leaf litter and feeding on bacteria. When food becomes scarce, some slime mold cells gather together and behave as a coordinated group. In cellular slime molds, individual cells can aggregate to form a multicellular-like structure that produces spores. Some cells become reproductive spores, while others form stalk-like structures that help lift and disperse those spores. This means some cells sacrifice their own reproduction to help other cells spread. Slime molds therefore provide a powerful example of how cooperation, cell signaling, and division of labor can evolve outside animals, plants, and fungi.

Figure 14. Slime molds. Slime molds can live as individual amoeboid cells but may aggregate under stressful conditions to form fruiting bodies. These life cycles show how cooperation and division of labor can evolve among eukaryotic cells.
Opisthokonta: Animals, Fungi, and Their Protist Relatives
Domain Eukarya: Opisthokonta
Opisthokonta is the major eukaryotic lineage that includes animals, fungi, and several unicellular or colonial relatives. Many opisthokonts are heterotrophs, meaning they obtain energy and carbon by consuming or absorbing organic matter. A key feature associated with the group is the presence of a single posterior flagellum in many motile cells. This can be seen in animal sperm cells and in the motile spores of some fungi, such as chytrids. In contrast, many other flagellated eukaryotes pull themselves forward with anterior flagella. Opisthokonta is important because it contains two of the most familiar multicellular kingdoms: Animalia and Fungi. These two groups evolved multicellularity independently and solve the problem of feeding in very different ways. Animals usually ingest food and digest it internally. Fungi usually digest externally by secreting enzymes and absorbing the released nutrients. Their shared ancestry reminds students that familiar kingdoms are nested within a much broader diversity of microbial eukaryotes.
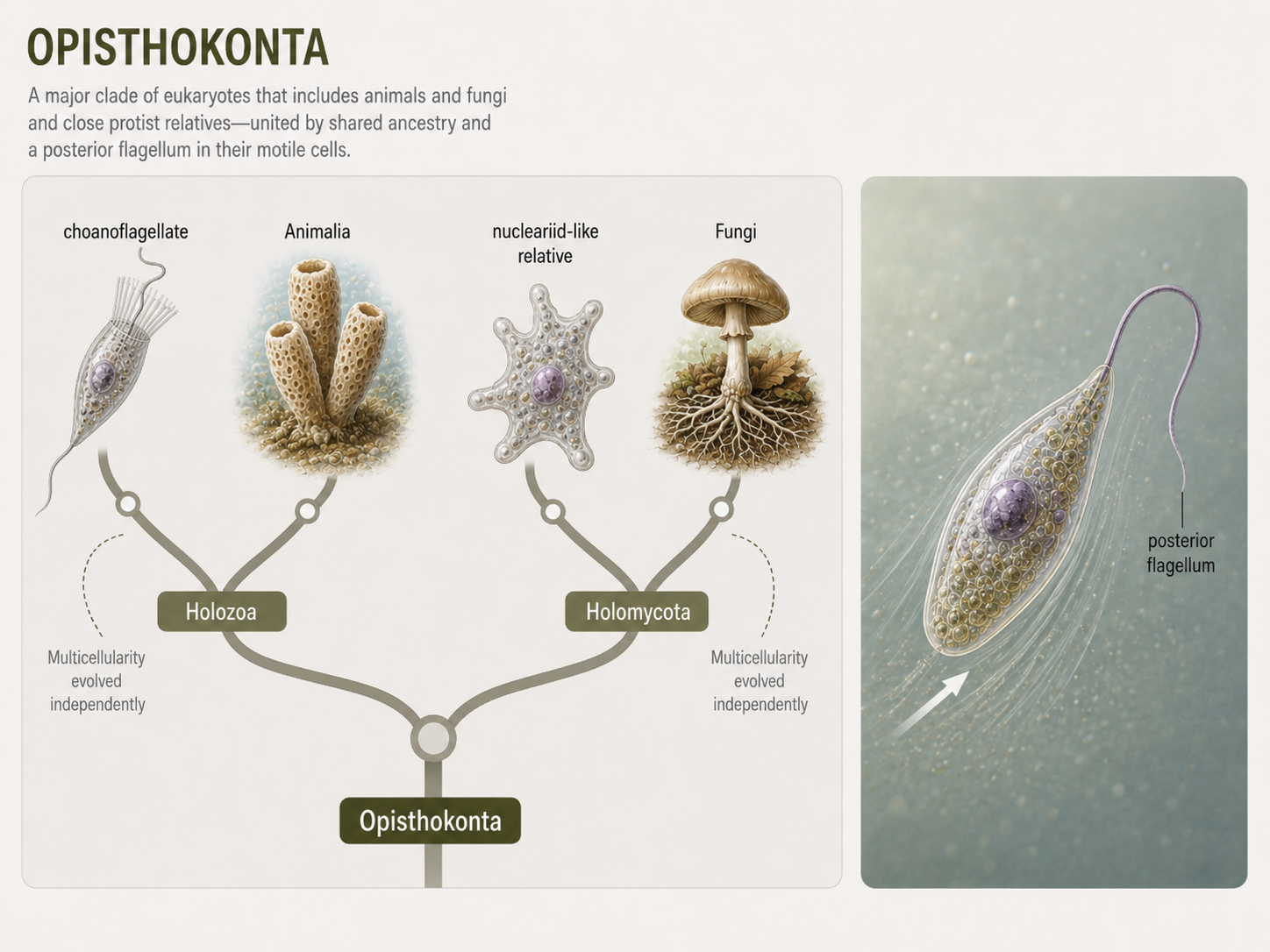
Figure 15. Opisthokonta. Opisthokonts include animals, fungi, and their close protist relatives. Many have cells with a single posterior flagellum, and the group contains two independently multicellular lineages with very different feeding strategies.
Holomycota: Fungi and Their Relatives
Domain Eukarya: Opisthokonta: Holomycota
Holomycota includes fungi and their close unicellular relatives, such as nucleariids. Fungi are best known for external digestion. Instead of swallowing food, fungi secrete digestive enzymes into their surroundings and absorb the smaller molecules that result. This feeding strategy allows fungi to decompose dead organisms, form mutualisms with plants, and act as parasites or pathogens in some species. Fungi are distinguished from many of their close protist relatives by the presence of a chitin-containing cell wall. Chitin is a tough polysaccharide that provides structural support. Fungal bodies are often built from thread-like filaments called hyphae, which grow through food sources and increase surface area for absorption. Nucleariids, by contrast, are amoeba-like unicellular relatives of fungi that use thin pseudopodia for movement and feeding. Comparing fungi with nucleariids helps show how multicellular fungal bodies evolved from unicellular opisthokont ancestors.
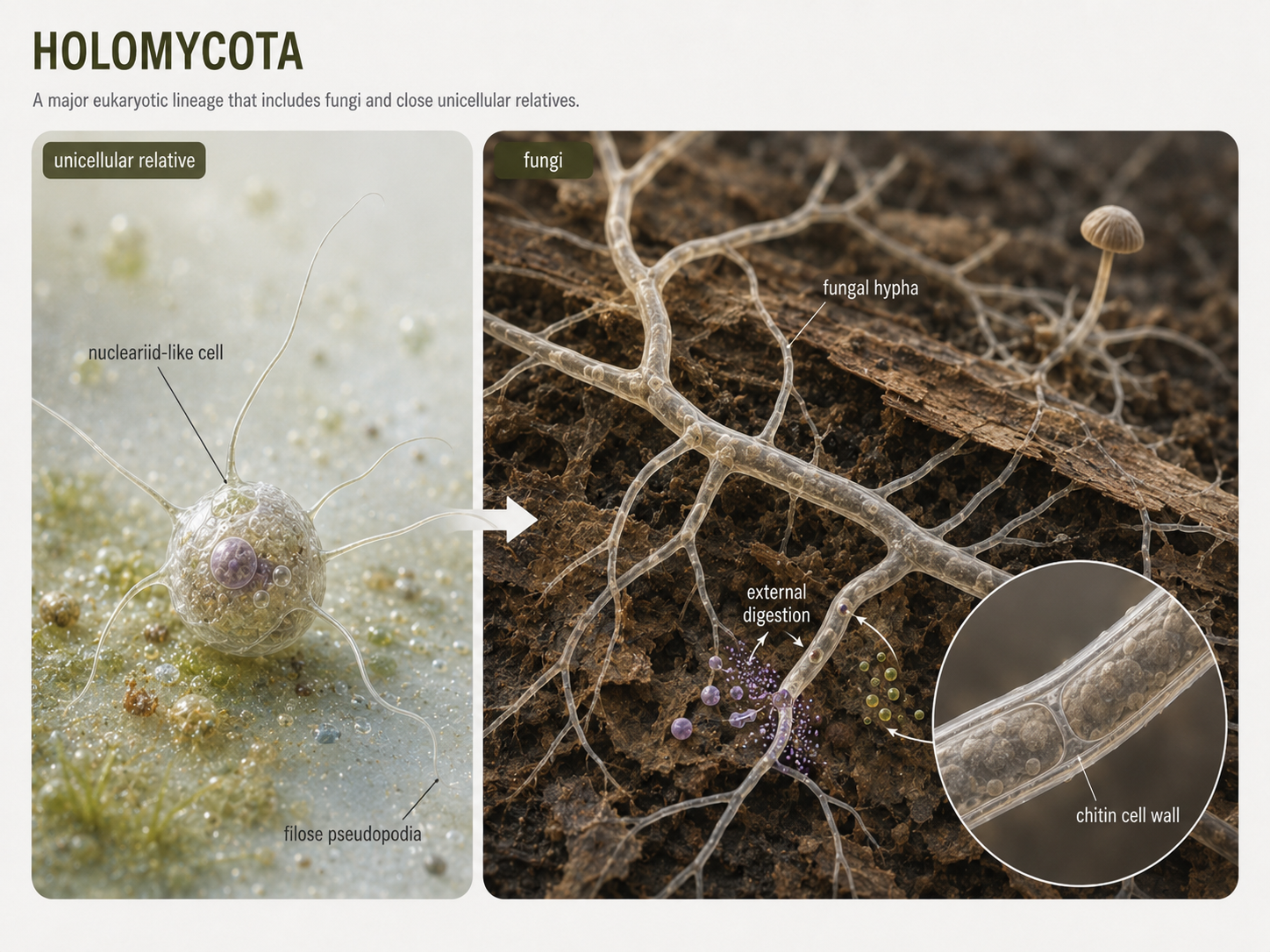
Figure 16. Holomycota. Holomycota includes fungi and close unicellular relatives. Fungi digest food externally and absorb nutrients, often using hyphae to grow through their food source. Their chitin-containing cell walls distinguish them from many close protist relatives.
Holozoa: Animals and Their Closest Relatives
Domain Eukarya: Opisthokonta: Holozoa
Holozoa includes animals and their closest unicellular and colonial relatives, including choanoflagellates. Animals are multicellular heterotrophs that ingest food and digest it internally. Choanoflagellates are not animals, but they are among the closest living relatives of animals and provide clues about how animal multicellularity evolved. They use a flagellum surrounded by a collar of microvilli to create water currents and capture bacteria. The connection between choanoflagellates and animals is especially clear in sponges. Sponges contain feeding cells called choanocytes that strongly resemble choanoflagellates in structure and function. This similarity supports the idea that early animals evolved from a choanoflagellate-like ancestor in which cells remained attached, communicated, and gradually specialized. Holozoa therefore links the origin of animals to a broader story of eukaryotic cell cooperation.
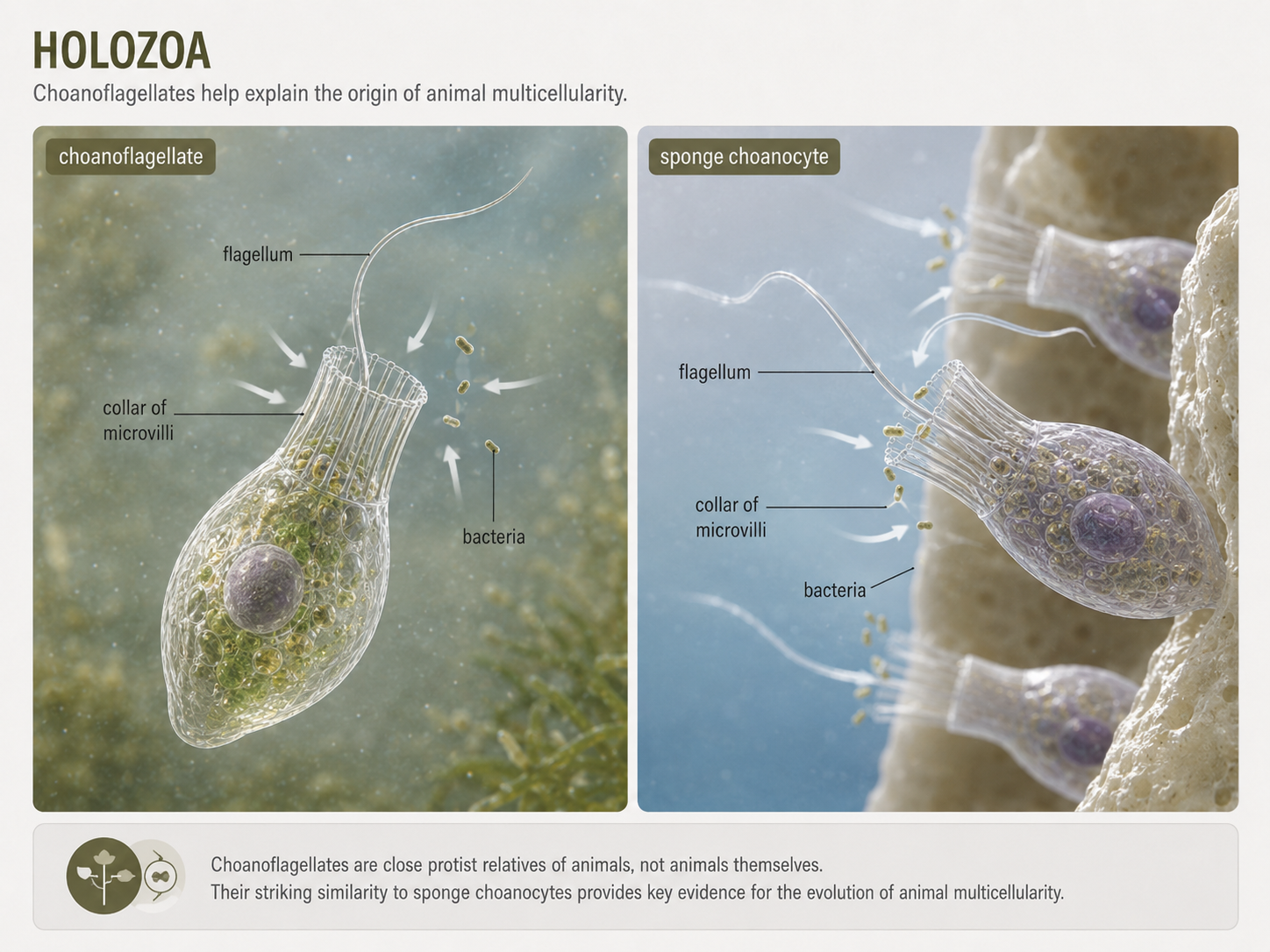
Figure 17. Holozoa. Holozoa includes animals and close relatives such as choanoflagellates. Choanoflagellates help explain the origin of animal multicellularity because their feeding cells resemble sponge choanocytes.
SAR: Stramenopiles, Alveolates, and Rhizarians
Domain Eukarya: SAR
SAR is a major eukaryotic grouping that includes Stramenopiles, Alveolates, and Rhizaria. This updated grouping replaces much of the older Chromoalveolata framework. SAR contains an enormous range of eukaryotes, including photosynthetic algae, microbial predators, parasites, shell-building plankton, kelps, diatoms, ciliates, dinoflagellates, foraminiferans, radiolarians, and oomycetes. SAR is difficult to summarize with one simple body trait because its members are extremely diverse. Some are photosynthetic because they contain plastids derived from red algae through secondary endosymbiosis. Others are heterotrophs or parasites, and some lineages have lost photosynthesis. SAR is best understood as a major eukaryotic branch that contains several ecologically important groups, especially in aquatic ecosystems. Many SAR organisms form plankton, drive marine food webs, build sediments, cause disease, or contribute to global carbon cycling.
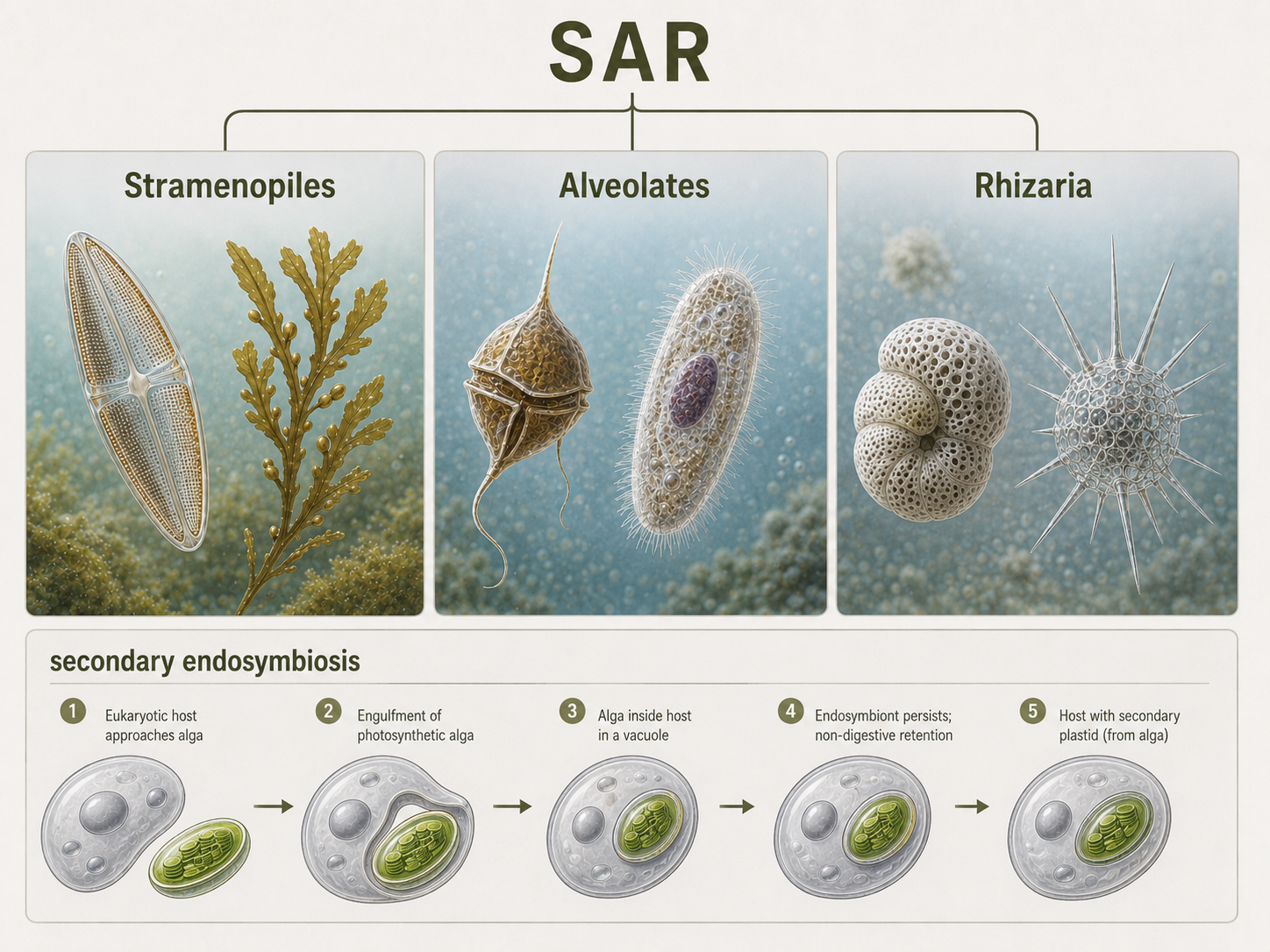
Figure 18. SAR. SAR includes Stramenopiles, Alveolates, and Rhizaria. This diverse eukaryotic group contains photosynthetic algae, microbial predators, parasites, and shell-building plankton that play major roles in aquatic ecosystems.
Rhizaria: Thread-Like Pseudopodia and Intricate Skeletons
Domain Eukarya: SAR: Rhizaria
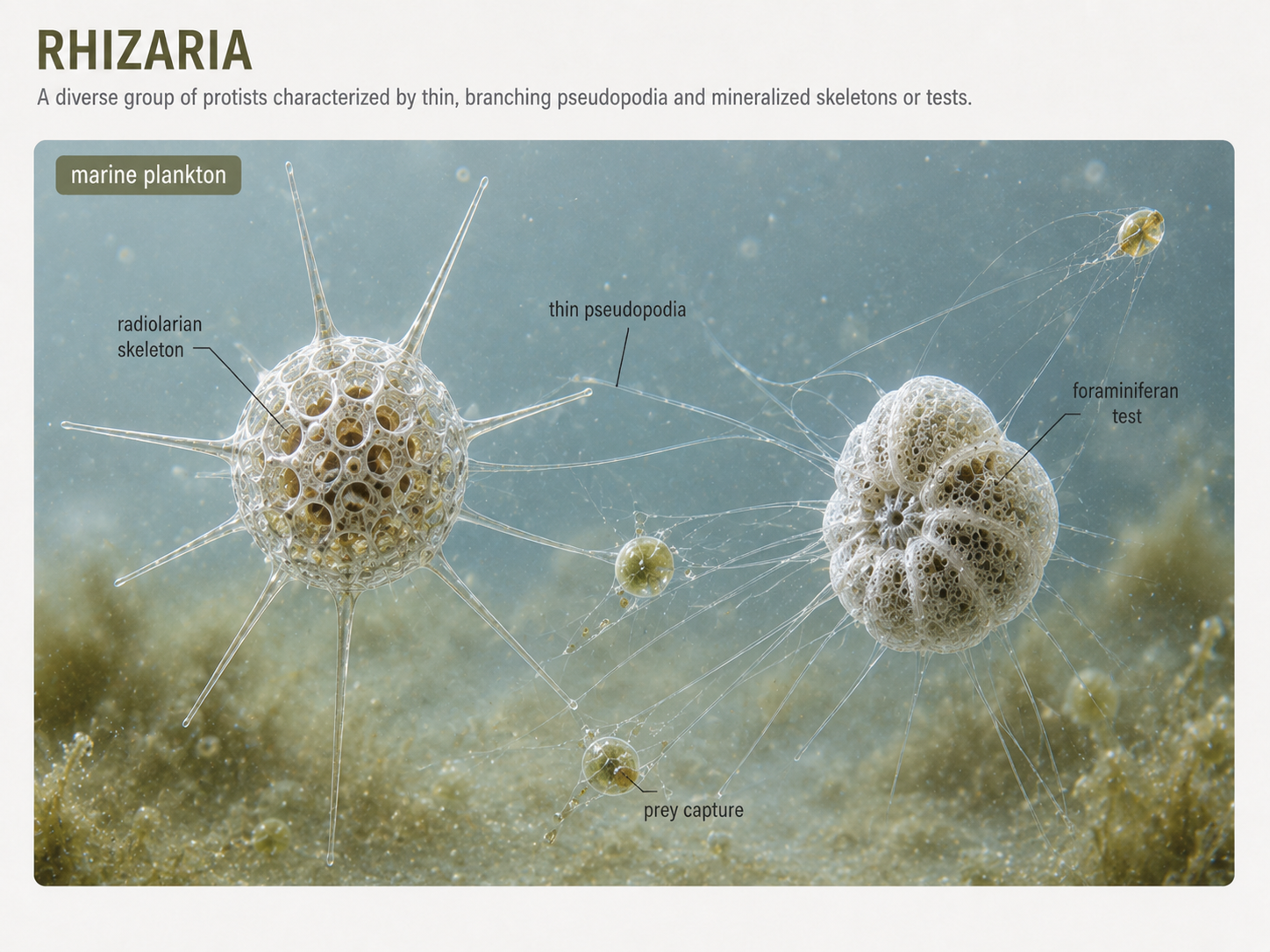
Rhizarians are mostly unicellular eukaryotes that often use thin, thread-like pseudopodia to capture food. These pseudopodia differ from the blunt lobose pseudopodia of many amoebozoans. Rhizarian pseudopodia can extend outward like fine nets or needles, increasing the cell’s ability to trap bacteria, algae, and other small prey. Many rhizarians are common in marine plankton and sediments. Radiolarians and foraminiferans are especially important rhizarians. Radiolarians often have delicate mineral skeletons and needle-like projections supported by microtubules. Foraminiferans usually have porous shells called tests through which pseudopodia extend. When these organisms die, their shells can accumulate on the seafloor and become part of the fossil record. Because foraminiferans shells preserve well and vary through time, they are useful in studies of ancient climates and ocean conditions.
Figure 19. Rhizaria. Rhizarians often use thin pseudopodia to capture prey. Radiolarians and foraminiferans are important marine plankton with mineralized skeletons or shells that can preserve in sediments and contribute to the fossil record.
Cercozoans and Chlorarachniophytes
Domain Eukarya: SAR: Rhizaria: Cercozoa
Cercozoans are a diverse group of rhizarians that often move and feed using thin pseudopodia. Many are heterotrophic predators of bacteria and other protists. Some also have flagella, showing that eukaryotic movement can combine multiple structures in the same organism. One especially interesting cercozoan group is the chlorarachniophytes. These organisms acquired photosynthesis through secondary endosymbiosis with a green alga. As a result, they can photosynthesize but can also feed on other organisms. Chlorarachniophytes are mixotrophs, combining autotrophic and heterotrophic nutrition. They also retain a nucleomorph, a tiny remnant of the engulfed green algal nucleus, providing strong evidence for secondary endosymbiosis.
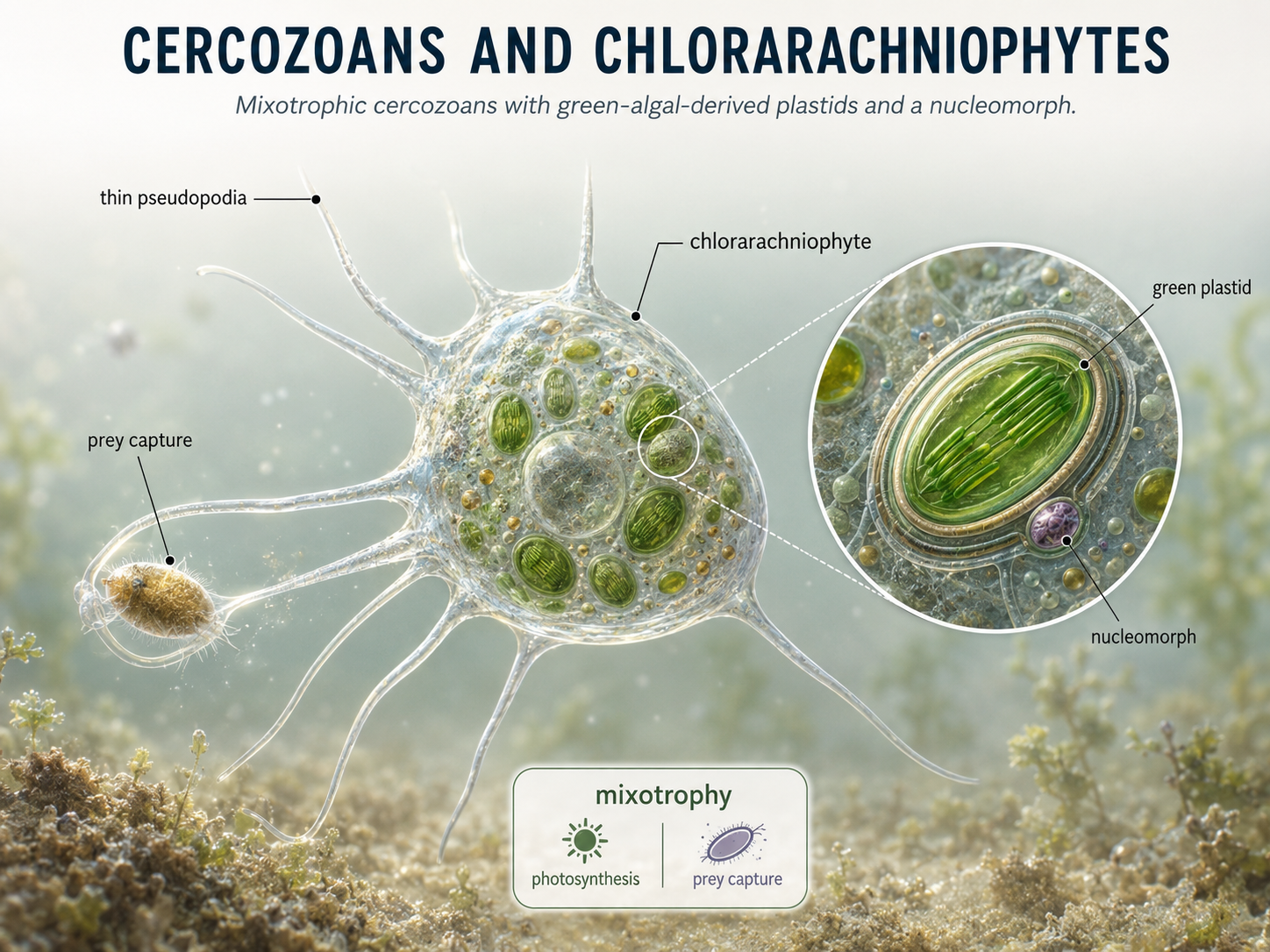
Figure 20. Cercozoans and chlorarachniophytes. Cercozoans are rhizarians that often use thin pseudopodia for feeding. Chlorarachniophytes are mixotrophic cercozoans with green algal-derived plastids and a nucleomorph, showing evidence of secondary endosymbiosis.
Alveolates: Membrane Sacs Beneath the Cell Surface
Domain Eukarya: SAR: Alveolata
Alveolates are a major SAR lineage defined by small membrane-bound sacs called alveoli located beneath the plasma membrane. These sacs may help support the cell surface, regulate cell shape, or contribute to buoyancy and protection, depending on the group. Alveolates include dinoflagellates, ciliates, and apicomplexans. These groups look very different from one another, but they share a common alveolate ancestry. Dinoflagellates are major components of marine plankton and include photosynthetic, heterotrophic, mixotrophic, and parasitic species. Ciliates are common microbial predators in freshwater and marine systems. Apicomplexans are specialized parasites, including Plasmodium, the organism that causes malaria. Together, alveolates show how one eukaryotic lineage can diversify into photosynthetic plankton, active predators, and highly specialized parasites.
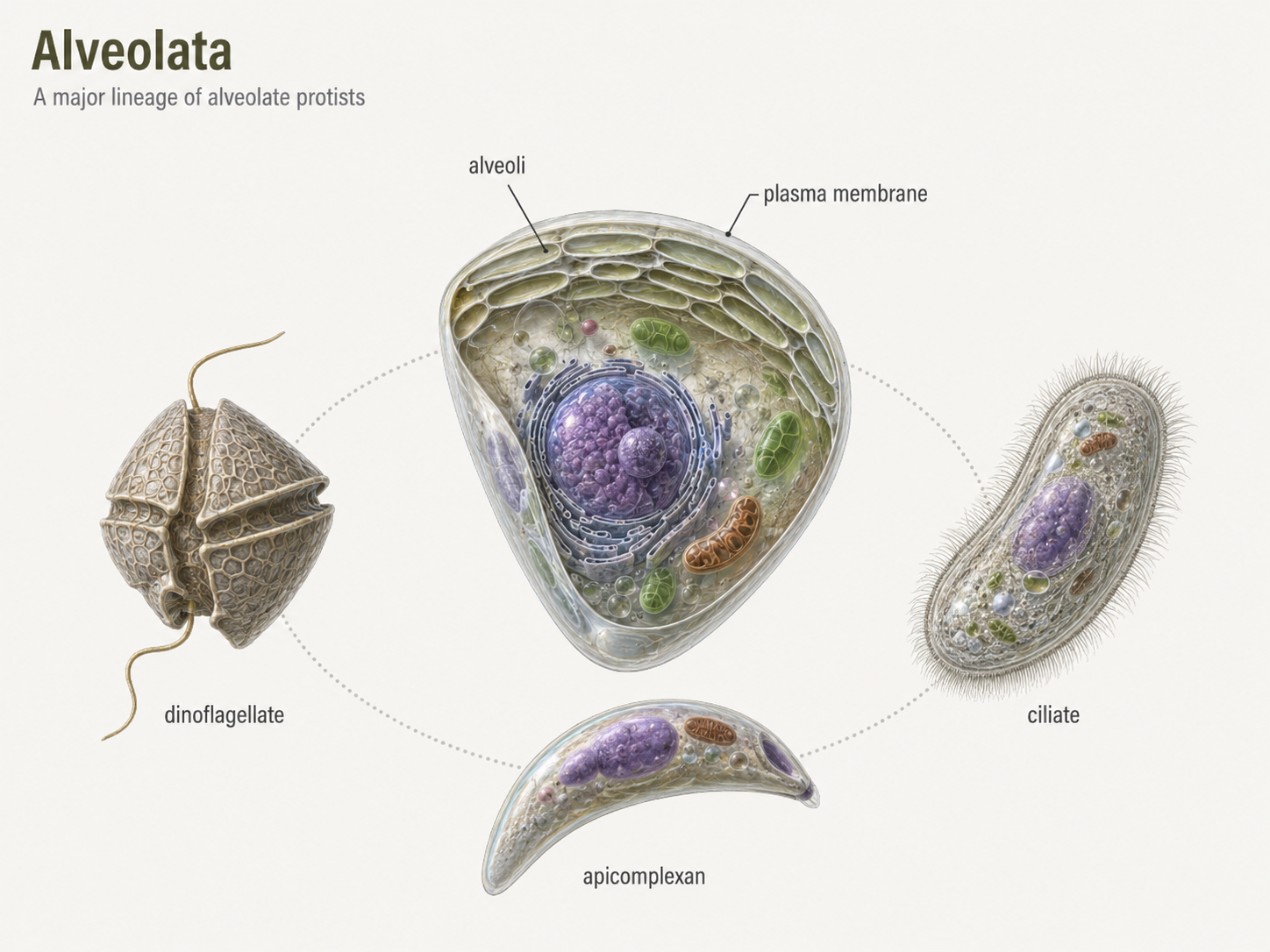
Figure 21. Alveolates. Alveolates are SAR eukaryotes with membrane sacs called alveoli beneath the cell surface. The group includes dinoflagellates, ciliates, and apicomplexans, which differ greatly in ecology and body organization.
Dinoflagellates: Photosynthesis, Flagella, and Blooms
Domain Eukarya: SAR: Alveolata: Dinoflagellata
Dinoflagellates are mostly unicellular alveolates that often have two flagella and, in many species, cellulose-reinforced plates around the cell. Some dinoflagellates are photosynthetic, while others are heterotrophic or mixotrophic. Many are important members of marine and freshwater plankton. Some dinoflagellates can undergo rapid population growth called blooms. When certain species bloom, they may produce toxins or create conditions that harm fish, shellfish, marine mammals, and humans. These harmful algal blooms are sometimes called red tides, although they are not always red and are not always linked directly to tides. Other dinoflagellates are beneficial, including the photosynthetic symbionts that live inside corals and help support coral reef ecosystems
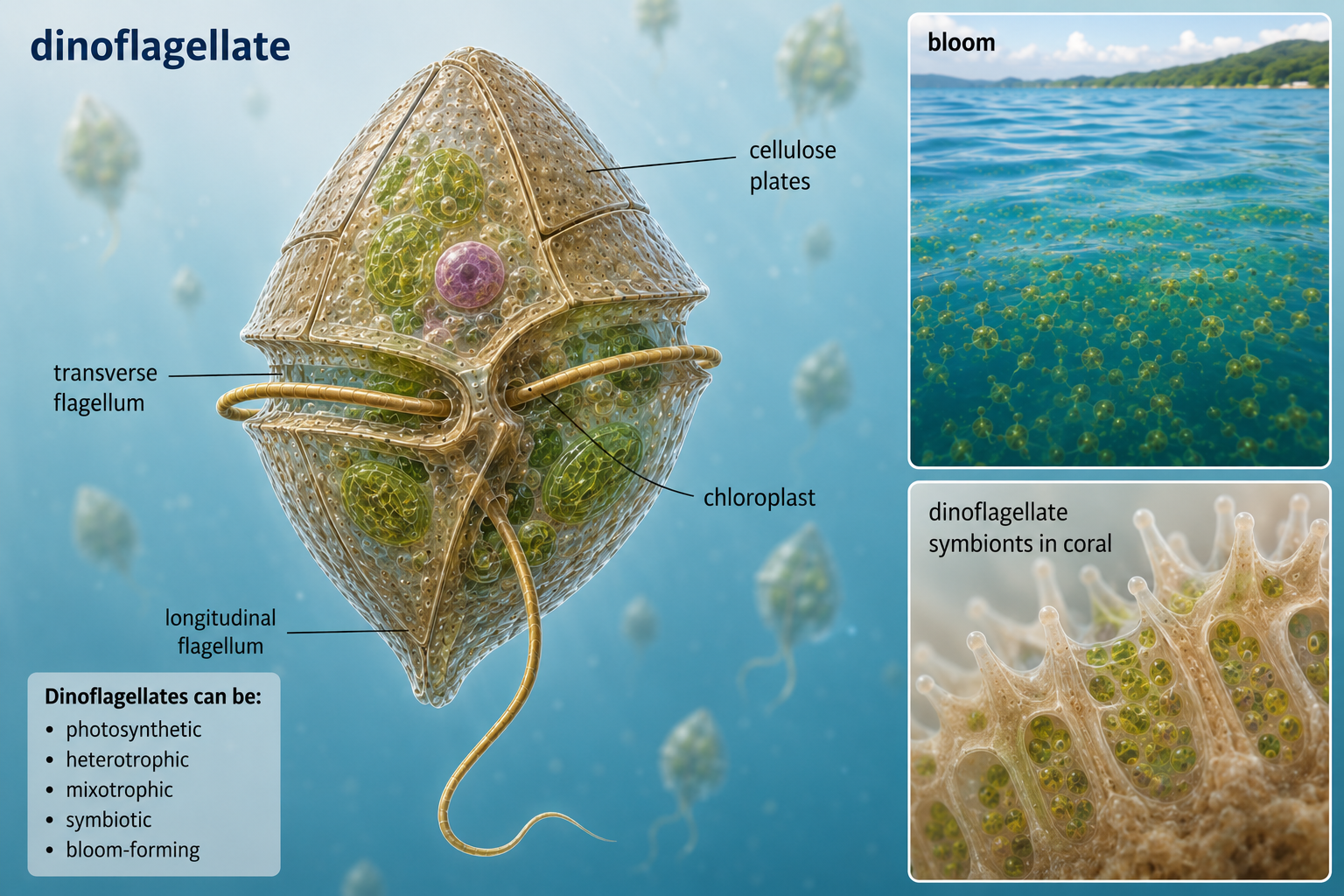
Figure 22. Dinoflagellates. Dinoflagellates are alveolates with two flagella and diverse nutritional strategies. Some are photosynthetic plankton, some form harmful algal blooms, and others live as important symbionts in corals.
Ciliates: Movement with Cilia
Domain Eukarya: SAR: Alveolata: Ciliata
Ciliates are alveolates covered with many short hair-like structures called cilia. These cilia beat in coordinated waves, allowing the cell to swim, crawl, create feeding currents, or move food toward an oral groove. Paramecium is a familiar example of a ciliate often studied in introductory biology. Ciliates are also unusual because they typically have two kinds of nuclei. The macronucleus controls everyday cell functions, including gene expression and metabolism. The micronucleus is involved in sexual reproduction and genetic exchange. This separation of nuclear function makes ciliates important examples of how complex cellular organization can evolve within a single cell. Like many protists, ciliates remind us that unicellular does not mean simple.
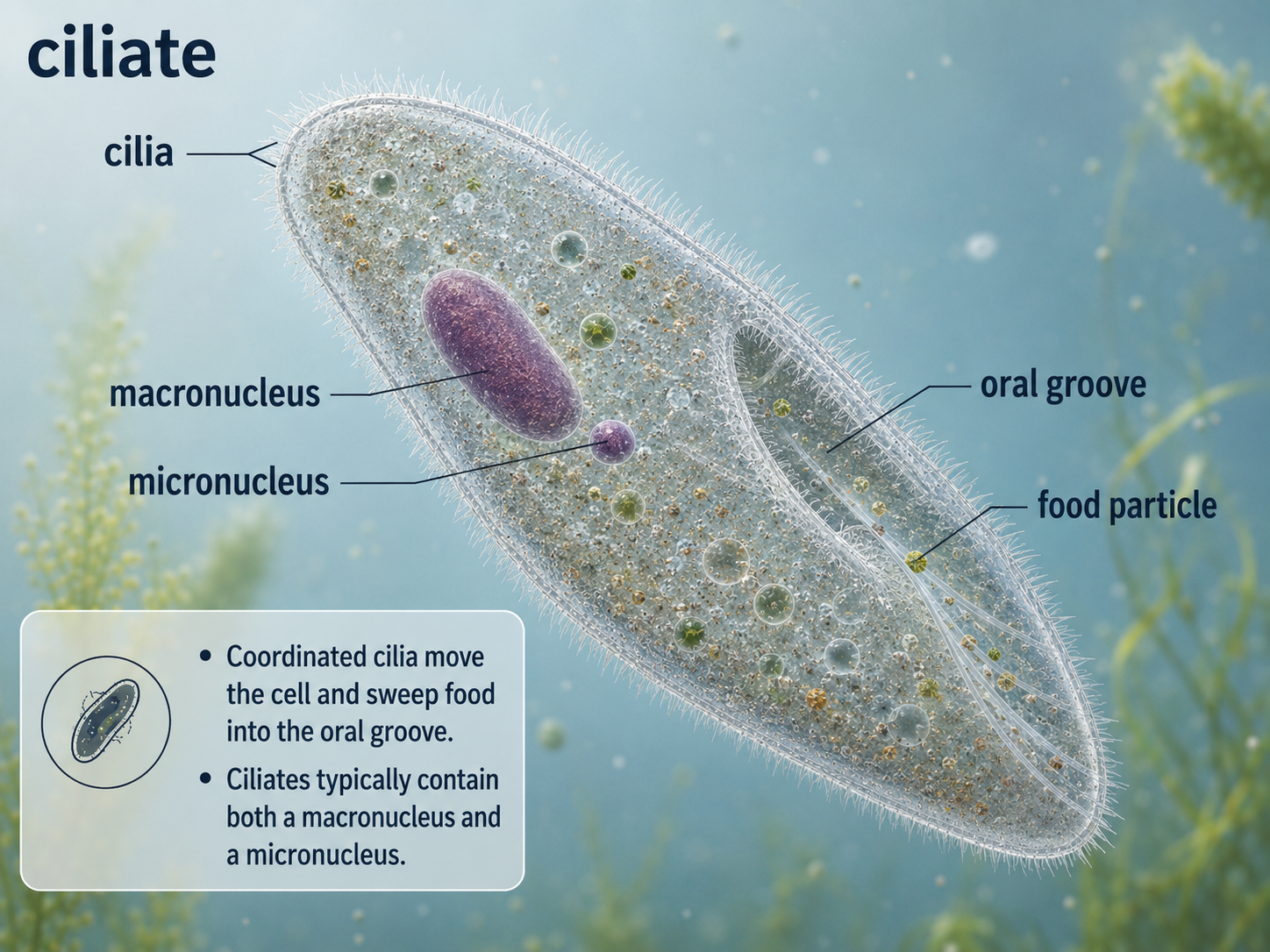
Figure 23. Ciliates. Ciliates use rows of cilia for movement and feeding. They usually contain a large macronucleus for everyday cell function and a small micronucleus involved in sexual reproduction.
Apicomplexans: Specialized Parasites
Domain Eukarya: SAR: Alveolata: Apicomplexa
Apicomplexans are parasitic alveolates that infect animals. They are named for an apical complex, a specialized set of structures used to enter host cells. Unlike free-living predators that engulf food from the environment, apicomplexans live inside host tissues or cells and obtain nutrients from their hosts. Plasmodium, the parasite that causes malaria, is one of the most important apicomplexans affecting humans. Although apicomplexans are not photosynthetic, many retain a plastid-derived organelle called an apicoplast. This organelle is evidence that their ancestors likely had a photosynthetic or plastid-containing past. Over evolutionary time, the lineage lost photosynthesis but retained a modified plastid that still performs essential metabolic functions. Apicomplexans therefore show how endosymbiotic organelles can be repurposed after photosynthesis is lost.
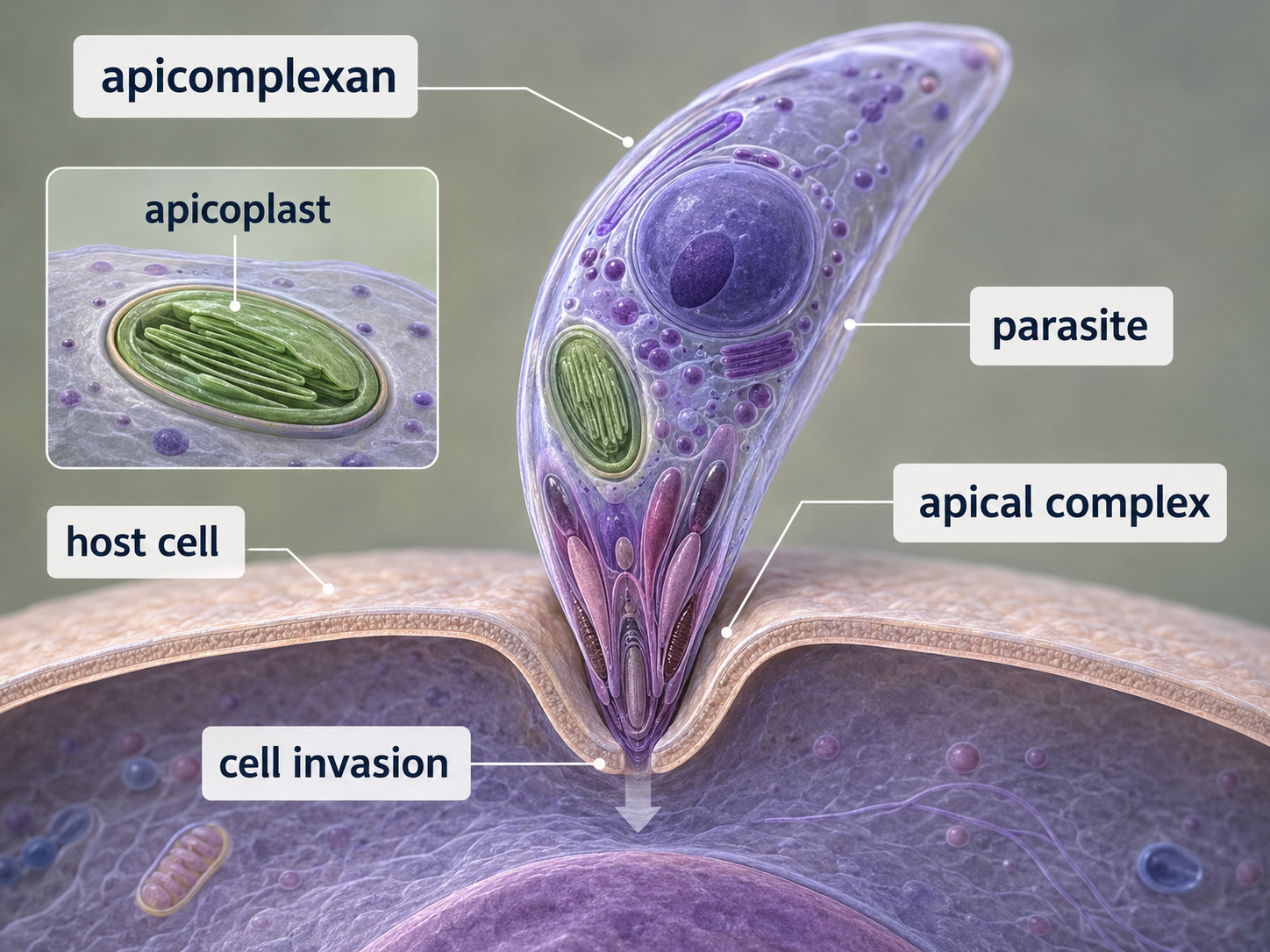
Figure 24. Apicomplexans. Apicomplexans are parasitic alveolates that use an apical complex to infect host cells. Many retain a modified plastid called an apicoplast, showing evidence of ancestry from plastid-containing eukaryotes.
Stramenopiles: Hairy Flagella, Diatoms, Brown Algae, and Oomycetes
Domain Eukarya: SAR: Stramenopiles
Stramenopiles are a major SAR lineage that includes diatoms, brown algae, and oomycetes. Many stramenopiles have two unequal flagella at some stage of life: one smooth flagellum and one “hairy” flagellum with fine projections. This trait gives the group its name and helps distinguish it from many other eukaryotes. Stramenopiles include both photosynthetic and nonphotosynthetic organisms. Photosynthetic stramenopiles, such as diatoms and brown algae, have plastids derived from secondary endosymbiosis involving a red alga. Other stramenopiles, such as oomycetes, have lost photosynthesis and live as decomposers or parasites. This diversity shows that once a lineage acquires a plastid, that plastid can be maintained, modified, reduced, or lost depending on ecological conditions and evolutionary history.

Figure 25. Stramenopiles. Stramenopiles include diatoms, brown algae, and oomycetes. Many have two unequal flagella, and photosynthetic members contain red algal-derived plastids from secondary endosymbiosis.
Diatoms: Glass-Walled Photosynthetic Plankton
Domain Eukarya: SAR: Stramenopiles: Diatoms
Diatoms are unicellular photosynthetic stramenopiles with cell walls made of silica, the same basic material found in glass. Their two-part silica walls fit together like a box and lid, forming intricate shapes that can be circular, elongated, or highly patterned. These walls provide protection and help diatoms resist predation and environmental stress. Diatoms are among the most important photosynthetic organisms in aquatic ecosystems. They are major primary producers in oceans, lakes, and rivers, converting carbon dioxide into organic molecules that support food webs. When diatoms die, some sink to the seafloor, carrying carbon with them and contributing to long-term carbon cycling. Their silica walls can also accumulate in sediments, preserving a record of past aquatic conditions.
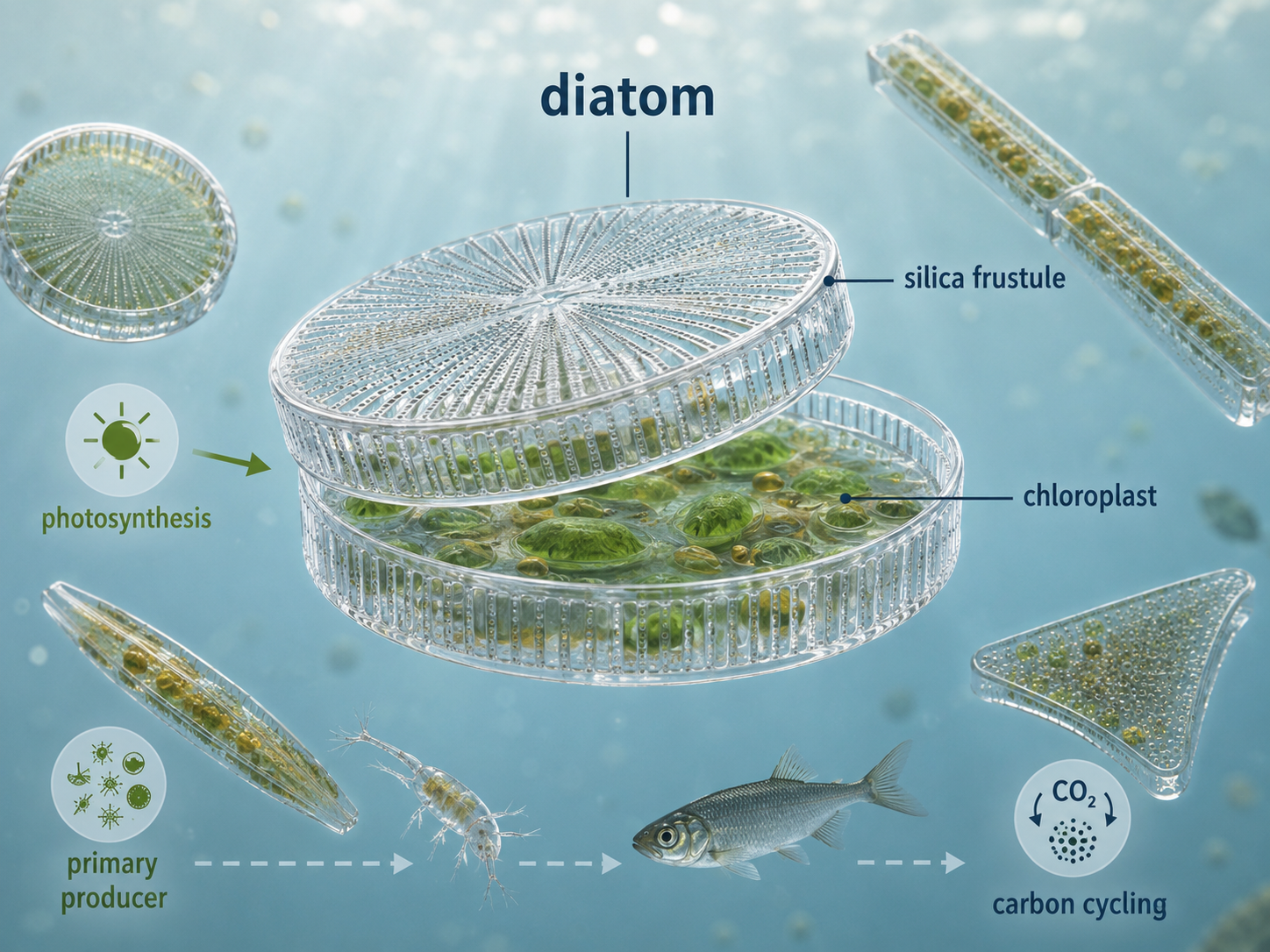
Figure 26. Diatoms. Diatoms are unicellular photosynthetic stramenopiles with silica cell walls. They are major aquatic primary producers and play important roles in food webs, carbon cycling, and sediment formation.
Brown Algae: Multicellularity in Stramenopiles
Domain Eukarya: SAR: Stramenopiles: Brown Algae
Brown algae are multicellular photosynthetic stramenopiles that include kelps and other large seaweeds. Although they may look plant-like, brown algae are not land plants. Their multicellularity evolved independently from the multicellularity of plants, animals, and fungi. This makes them an important example of repeated evolution of large multicellular bodies in Eukarya. Many brown algae have three major body regions. The holdfast anchors the organism to rocks or other surfaces. The stipe provides support, similar in appearance to a stem. The blades provide broad photosynthetic surfaces, similar in appearance to leaves. These structures are analogous to plant organs, meaning they perform similar functions but did not evolve from the same ancestral structures. Large kelps can form underwater forests that provide habitat, food, and shelter for many marine organisms.
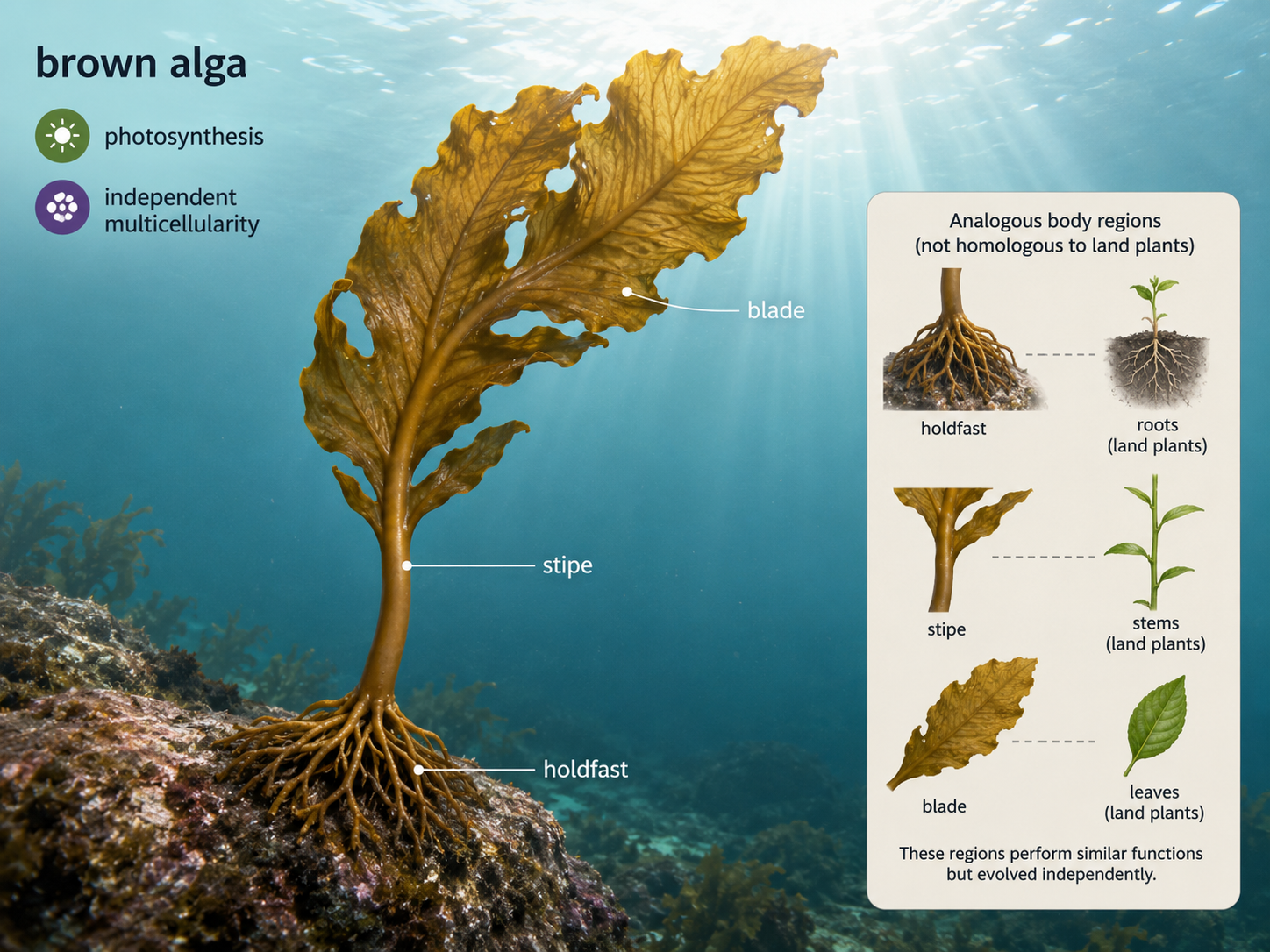
Figure 27. Brown algae. Brown algae are multicellular stramenopiles with plant-like structures such as holdfasts, stipes, and blades. These structures resemble plant organs in function but evolved independently.
Oomycetes: Fungus-Like Stramenopiles
Domain Eukarya: SAR: Stramenopiles: Oomycetes
Oomycetes include water molds, downy mildews, and several important plant pathogens. They often resemble fungi because they grow as filamentous bodies and absorb nutrients from their surroundings. However, oomycetes are not fungi. They belong to the stramenopiles, while true fungi belong to Opisthokonta. Their fungus-like form is an example of convergent evolution, in which unrelated groups evolve similar traits because they occupy similar ecological roles. Many oomycetes decompose dead organic matter in aquatic or moist environments. Others are parasites of plants and animals. Some plant-pathogenic oomycetes have had major effects on agriculture and history. Their ancestors likely had plastid-containing relatives, but modern oomycetes do not photosynthesize. Like apicomplexans, they show that photosynthetic ancestry can leave traces even in lineages that later become nonphotosynthetic parasites or decomposers.
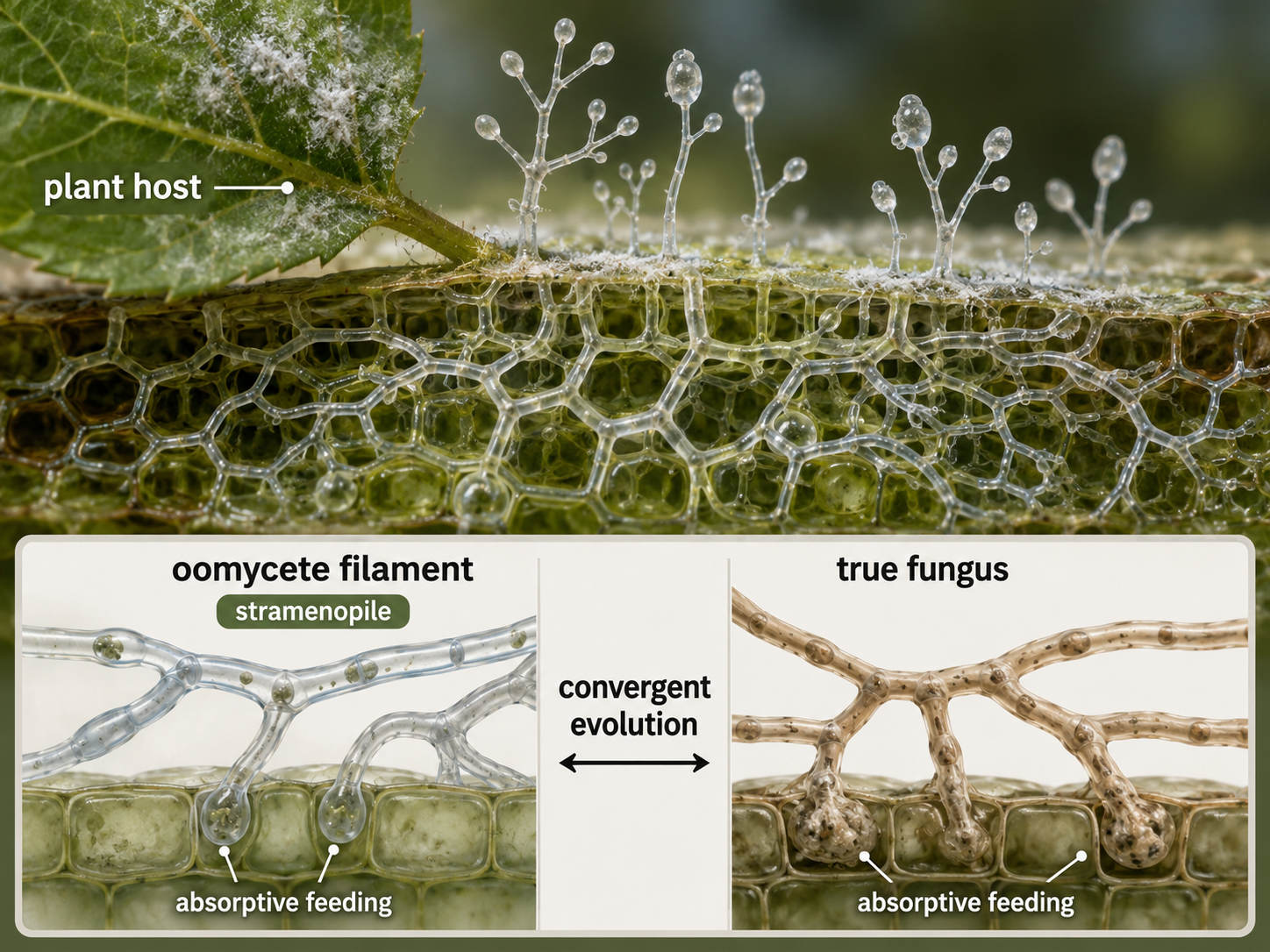
Figure 28. Oomycetes. Oomycetes are fungus-like stramenopiles that absorb nutrients and often grow as filaments. They are not true fungi; their similarity to fungi reflects convergent evolution.
Archaeplastida: Primary Plastids and the Lineage of Plants
Domain Eukarya: Archaeplastida
Archaeplastida includes glaucophyte algae, red algae, green algae, and land plants. The major shared feature of this group is the primary plastid, a chloroplast descended directly from an engulfed cyanobacterium. Unlike plastids that spread through secondary endosymbiosis, Archaeplastida plastids trace back to the original primary endosymbiotic event. Red algae are mostly aquatic photosynthetic eukaryotes, many of which live in marine environments. Green algae include unicellular, colonial, and multicellular forms, and one branch of green algae gave rise to land plants. Glaucophyte algae are a smaller group but are important because they preserve features that help scientists understand early plastid evolution. Land plants later evolved from green algal relatives and transformed terrestrial environments. Archaeplastida is important because it links the origin of chloroplasts to the evolution of photosynthetic eukaryotes and land plants. It also helps explain why plants are not isolated from protist diversity. Plants are one branch within a larger eukaryotic lineage that includes many algal forms.
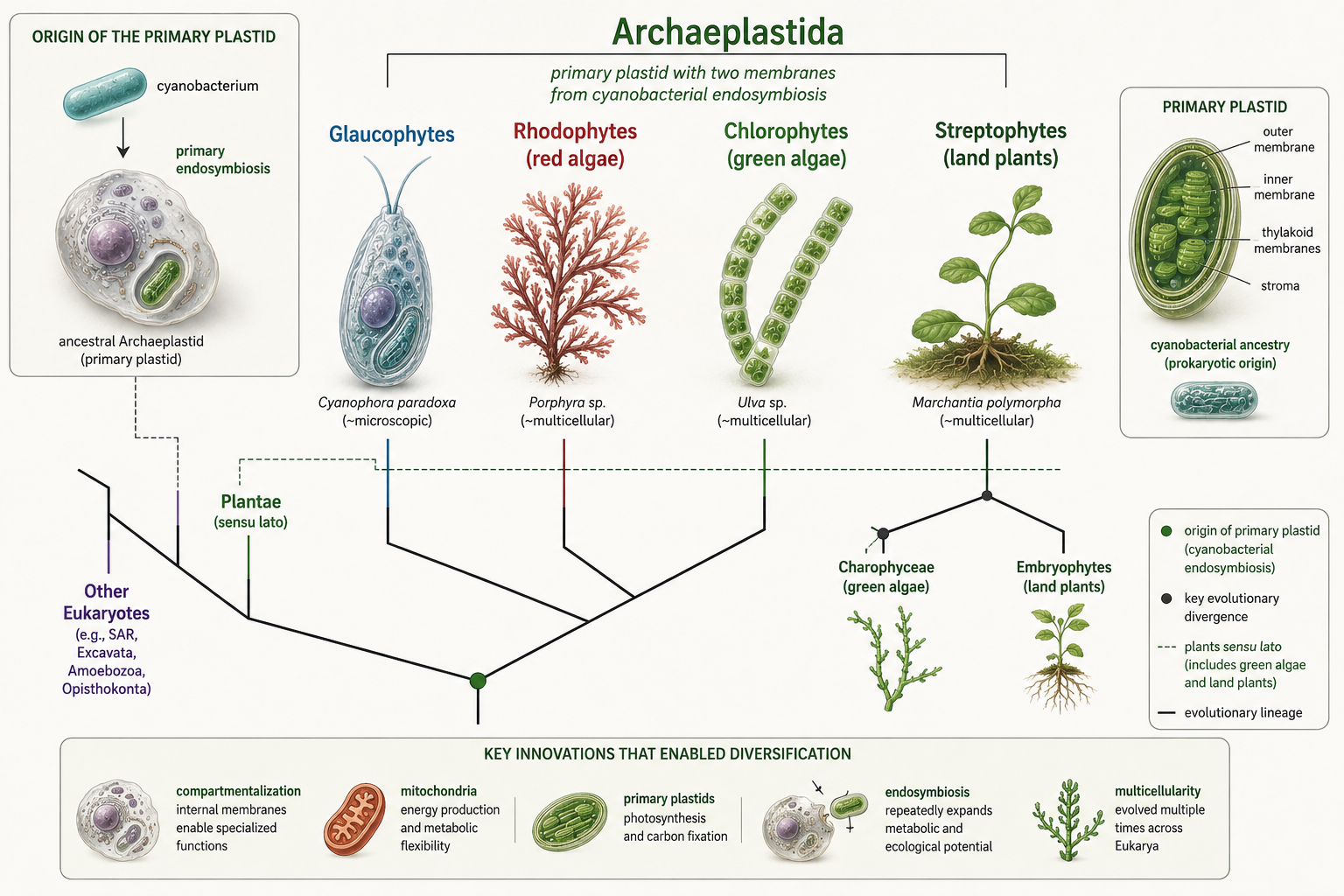
Figure 29. Archaeplastida. Archaeplastida includes glaucophytes, red algae, green algae, and land plants. These organisms share primary plastids descended from an ancient cyanobacterial endosymbiont.
Why Eukarya Matters
Domain Eukarya
Eukarya includes the organisms most familiar to humans, including animals, plants, and fungi, but its deepest diversity is microbial. Eukaryotes produce oxygen, form the base of aquatic food webs, decompose organic matter, cause diseases, build reefs and sediments, form forests on land and underwater, and shape global nutrient cycles. Many of the most important events in eukaryotic history were not the origin of large bodies, but the origin of complex cells. The story of Eukarya is a story of compartments, partnerships, and repeated innovation. The nucleus changed how cells controlled genes. The endomembrane system organized the inside of the cell. Mitochondria transformed cellular energy. Chloroplasts brought photosynthesis into eukaryotic cells. Secondary endosymbiosis moved photosynthesis into new lineages. Multicellularity evolved repeatedly in animals, fungi, plants, brown algae, red algae, green algae, and slime mold life cycles. These patterns show that evolution often works by modifying existing structures, forming new partnerships, and reusing old tools in new ways.
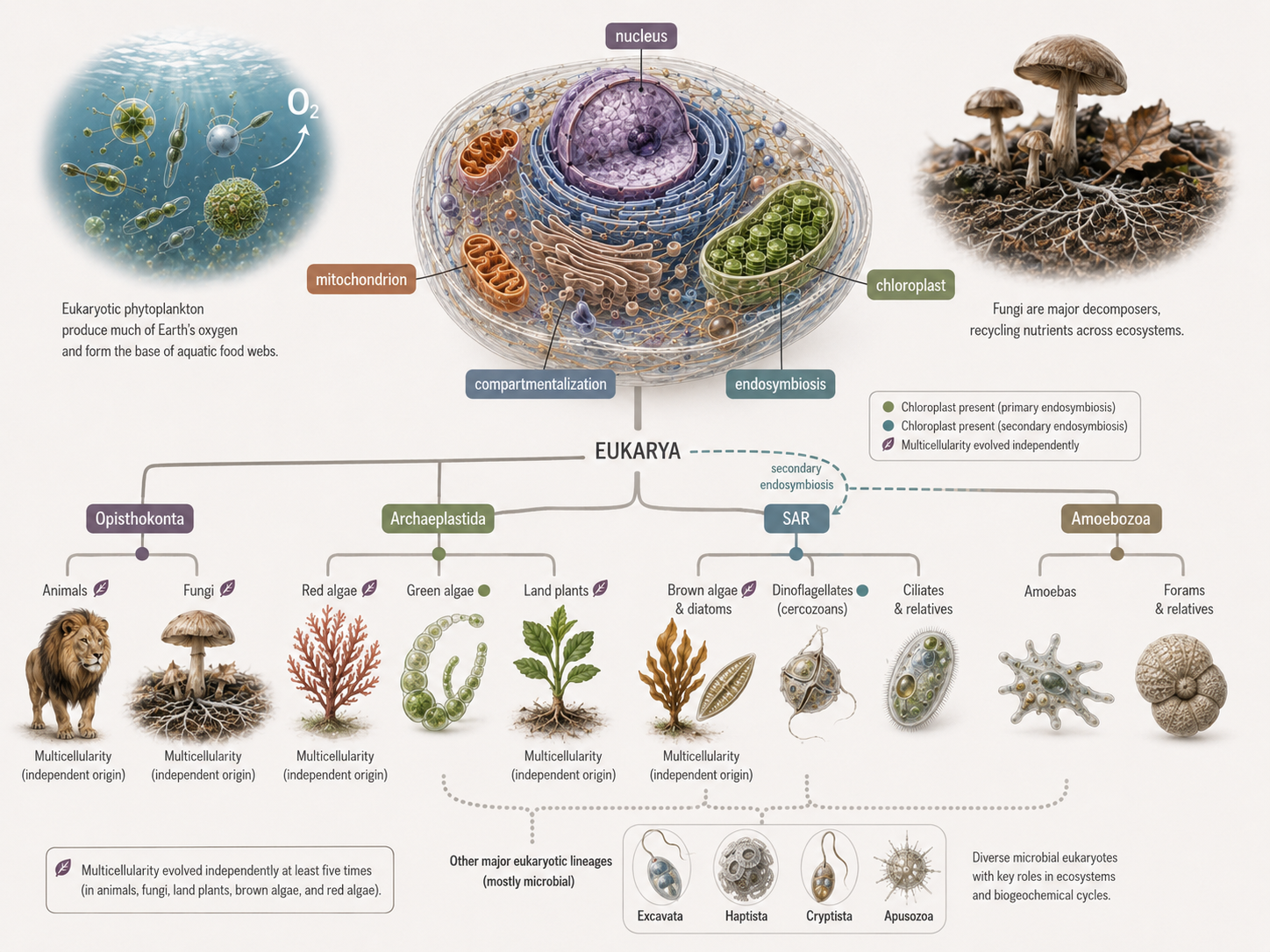
Figure 30. The evolution of Eukarya. Eukaryotic diversity was shaped by the origin of the nucleus, endomembrane system, mitochondria, chloroplasts, secondary endosymbiosis, and repeated multicellularity. These innovations produced the major eukaryotic lineages that shape modern ecosystems.