
Chapter: Prokaryotes
Bacteria and Archaea are two of the major lineages of cellular life. They are microscopic, ancient, diverse, and extraordinarily successful. They live in oceans, soils, sediments, hot springs, deep-sea vents, the atmosphere, glaciers, animal digestive systems, plant tissues, and nearly every other environment that has been studied. Although they are often invisible to us, they are among the most important organisms on Earth. Contemporary biologists often organize life into three main branches, or Domains: Bacteria, Archaea, and Eukarya. The three domain model grew out of molecular comparisons, especially ribosomal RNA studies associated with Carl Woese and colleagues, which showed that Archaea are deeply different from Bacteria. The three-domain model remains an important teaching framework, but current phylogenomic research often places Eukarya as closely related to, or emerging from within, Archaea, especially near the Asgard archaea. This means that the traditional three-domain diagram is useful, but it is not the final word on the deepest branches of life. This chapter focuses on Bacteria and Archaea. Together, they are often called prokaryotes, but that word needs to be used carefully.
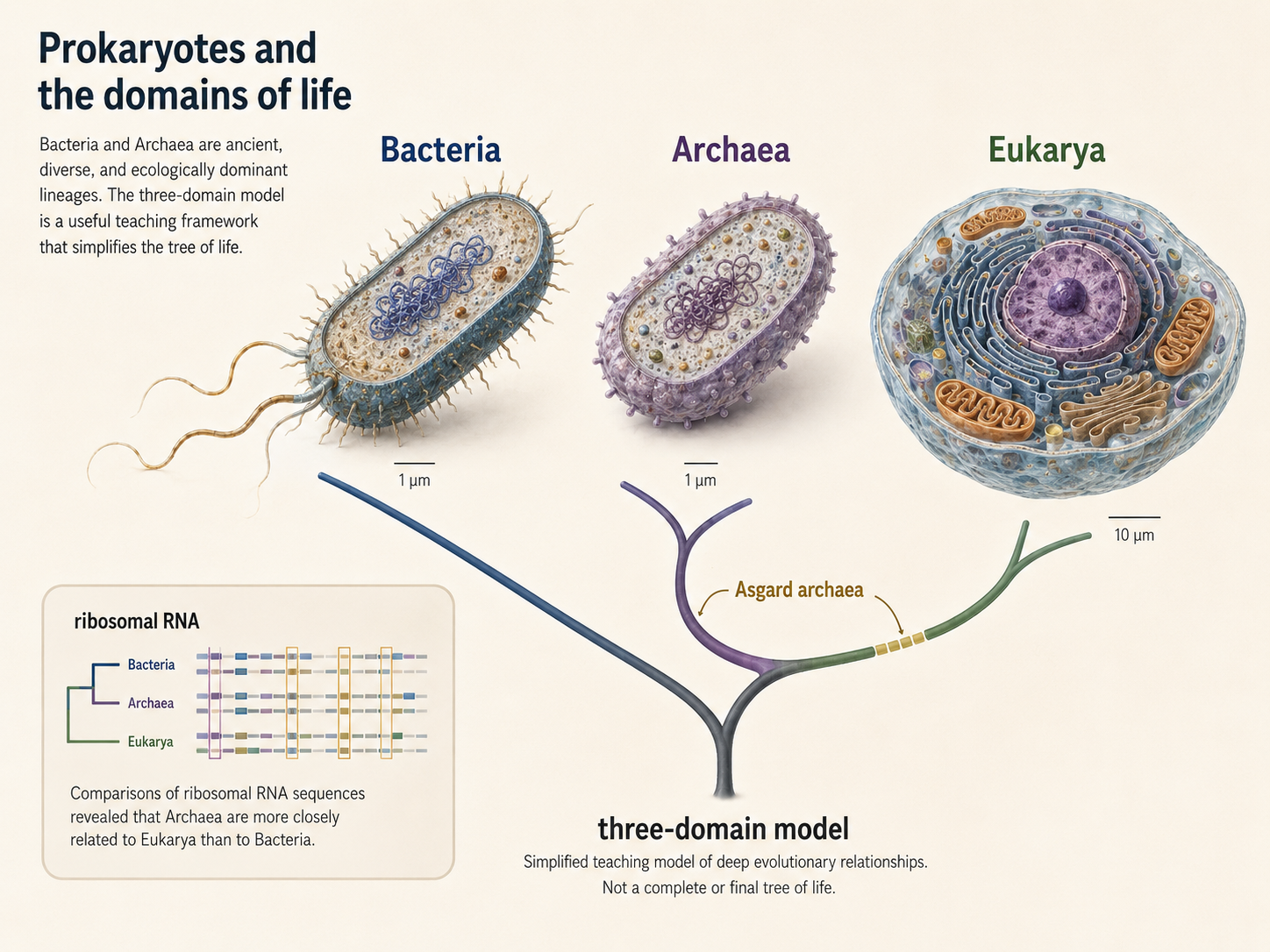
Figure 1. Bacteria, Archaea, and the Domains of Life. Bacteria and Archaea are ancient, diverse prokaryotic lineages, while Eukarya is closely connected to archaeal ancestry and bacterial endosymbiosis. The three-domain model remains useful for teaching, but modern evidence shows that the deepest relationships among life are more complex than a simple three-branch diagram.
What is a Prokaryote?
“Prokaryote” is useful for cell biology, but it does not represent one clean evolutionary clade. Prokaryotes are small, unicellular cells that lack a nucleus and lack organelles, typical of eukaryotic cells. Prokaryotic organisms belong to two Domains: Bacteria and Archaea. Prokaryotes is not a formal domain of life. This distinction matters because classification should reflect evolutionary relationships. However, Bacteria and Archaea are often grouped together as ‘prokaryotes’ due to several broad cell features they share. Members of both of these domains lack a nucleus, lack large membrane-bound organelles, and are usually much smaller than eukaryotic cells. Their DNA is found in a region called the nucleoid, rather than inside a nuclear envelope. Unlike a nucleus, the nucleoid is not enclosed by a membrane, so the DNA sits directly in the cytoplasm. Bacteria and Archaea both have one DNA molecule, often circular, and many also carry smaller DNA molecules called plasmids. These similarities make the word prokaryote useful in an introductory biology course. It allows us to compare prokaryotic cells with eukaryotic cells. However, Bacteria and Archaea are not simply two versions of the same thing. They differ in major molecular features, including membrane chemistry, cell wall chemistry, gene expression systems, and evolutionary history.
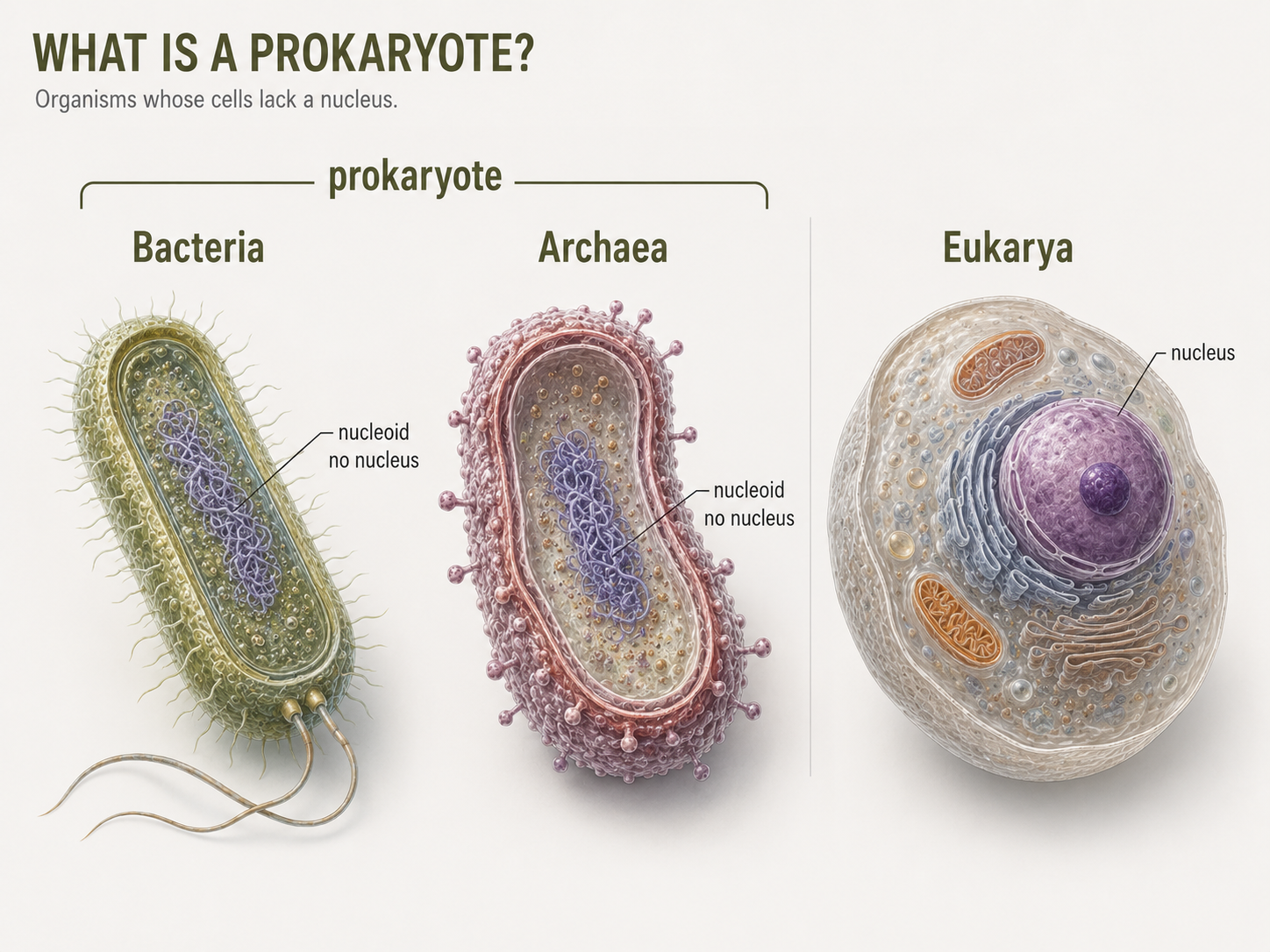
Figure 2. What Is a Prokaryote? “Prokaryote” describes cells that lack a membrane-bound nucleus, but it is not a formal domain of life. Bacteria and Archaea are both prokaryotic in cell structure, yet they are separate evolutionary lineages with important molecular differences.
Characteristics of Prokaryotic Cells
Most prokaryotic cells are very small. Their small size gives them a high surface-area-to-volume ratio, which helps them efficiently exchange materials quickly with the environment.This helps explain why many bacteria and archaea can grow and reproduce quickly when conditions are favorable. A typical prokaryotic cell includes a plasma membrane, cytoplasm, ribosomes, DNA in a nucleoid region, and often a cell wall. Many also have external structures such as capsules, pili, fimbriae, flagella, or archaella. These structures help cells survive, attach, move, exchange DNA, or interact with other organisms. Prokaryotes do not have a membrane-bound nucleus. They also lack the complex endomembrane system typical of eukaryotic cells, such as the endoplasmic reticulum and Golgi apparatus. However, it is not accurate to say that all prokaryotic cells are completely simple inside. Some bacteria have internal membranes, and some have protein-based compartments called bacterial microcompartments that organize specific metabolic reactions. These structures are not the same as eukaryotic organelles, but they show that prokaryotic cells can be more organized than used to think.
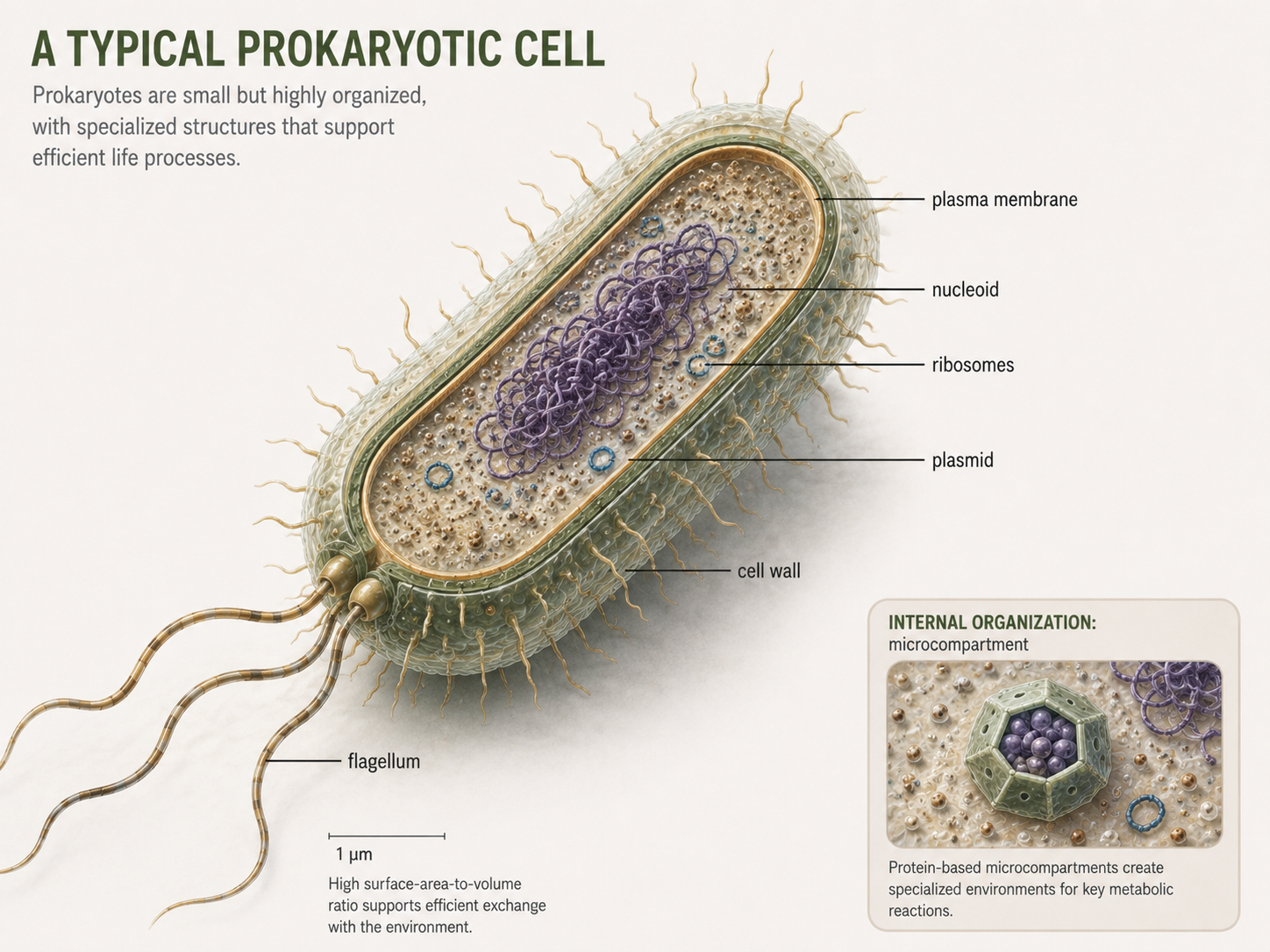
Figure 3. Characteristics of Prokaryotic Cells. Prokaryotic cells usually contain a plasma membrane, cytoplasm, ribosomes, DNA in a nucleoid region, and often a cell wall and external structures. Although they lack a nucleus and typical eukaryotic organelles, many prokaryotes have organized internal and surface features that support survival, movement, attachment, and reproduction.
The Prokaryotic Genome
The nucleoid is the region of a prokaryotic cell where most of the genetic material is located. Unlike the eukaryotic nucleus, the nucleoid is not surrounded by a membrane. Because prokaryotes do not have a nucleus, transcription and translation can occur in the same general cellular space. In eukaryotes, transcription, the process of using DNA to make mRNA, occurs inside the nucleus. The mRNA then leaves the nucleus and travels to a ribosome in the cytoplasm, where translation, the process of using mRNA to make a protein, occurs. In many bacteria, ribosomes can begin translating mRNA before transcription has fully finished. Most prokaryotes have one main chromosome that contains the genes needed for basic cell life. This chromosome is usually circular, but not always. Some prokaryotes have linear chromosomes or more than one chromosome. Many prokaryotes also have plasmids, which are smaller pieces of DNA separate from the main chromosome. Plasmids usually carry helpful but nonessential genes, such as genes for antibiotic resistance. The main chromosome is required for survival, while plasmids are extra DNA that can give the cell an advantage in certain environments.
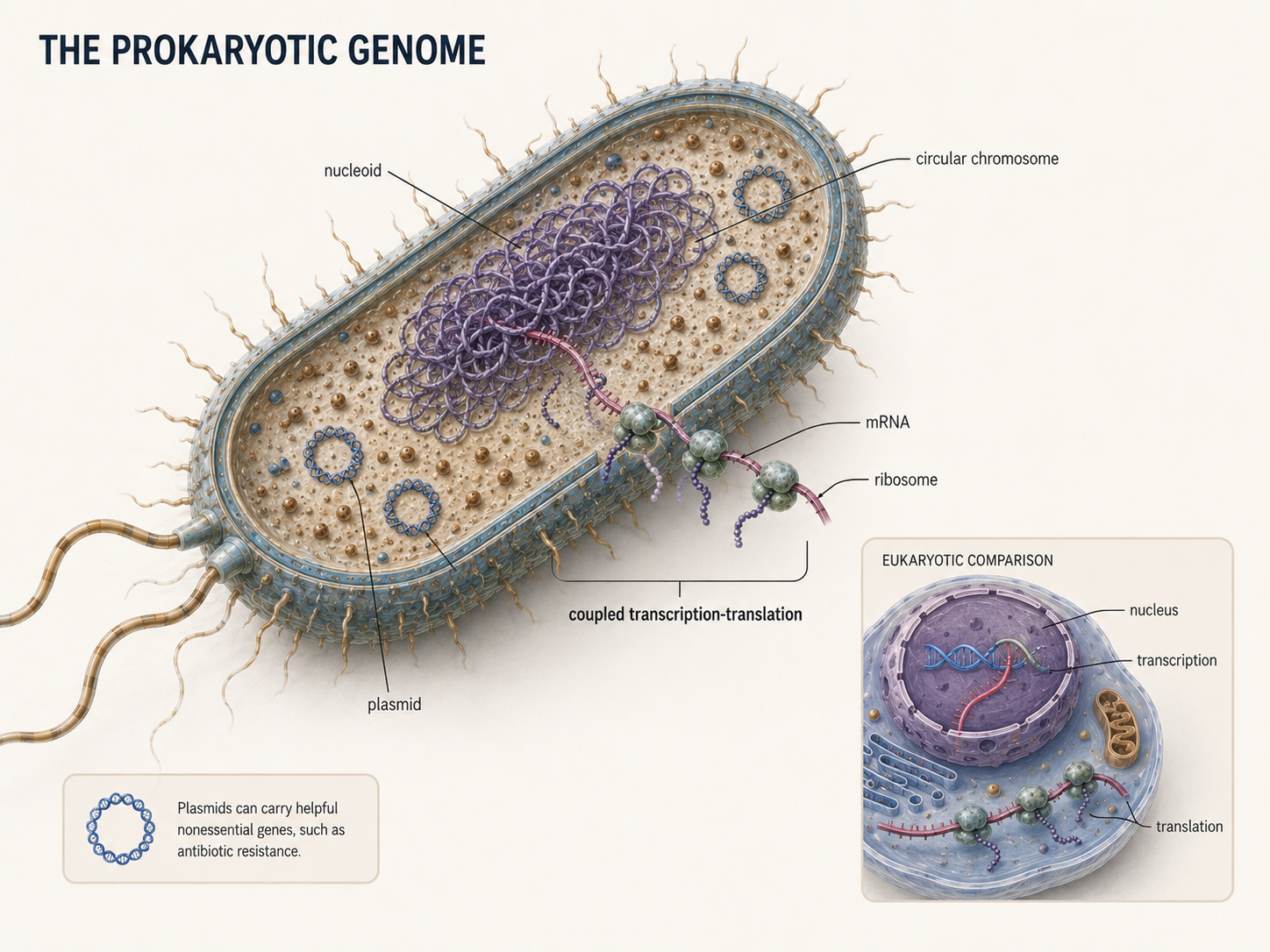
Figure 4. The Prokaryotic Genome. Most prokaryotes carry their main genetic material in a nucleoid region that is not surrounded by a nuclear envelope. Many also contain plasmids, which are smaller DNA molecules that may carry useful genes such as antibiotic resistance.
Horizontal Gene Transfer
Prokaryotes can evolve quickly because many of them reproduce rapidly. When a useful mutation appears, it can spread through a population in a short amount of time. However, mutations are not the only way prokaryotes gain genetic variation. They can also acquire genes from other organisms through horizontal gene transfer, which moves DNA sideways between cells instead of from parent to offspring. There are three major forms of horizontal gene transfer. In transformation, a cell simply takes up DNA from its environment. In transduction, viruses carry DNA from one cell to another, much the way they do in humans (i.e. HIV). In conjugation, DNA moves directly between cells, often through a connection junction. These processes allow prokaryotes to gain new traits quickly, including traits that may help them survive in changing environments. Horizontal gene transfer also makes prokaryotic evolution harder to show as a simple branching tree. In many organisms, genes mainly pass from parent to offspring, forming a familiar pattern known as vertical inheritance. In prokaryotes, genes do not only move from parent cells to offspring cells. They can also move sideways between different lineages through horizontal gene transfer. Because of this, prokaryotic evolution does not always look like a clean branching tree. It can look more like a network, with genes moving across branches as well as down from ancestors to descendants.
Figure 5. Horizontal Gene Transfer. Prokaryotes can gain genetic variation through transformation, transduction, and conjugation. These processes move DNA between cells and can spread traits such as antibiotic resistance across populations and lineages.
Reproduction by Binary Fission
Many bacteria and archaea reproduce by binary fission, a form of asexual reproduction in which one cell divides into two new cells. During binary fission, the cell first undergoes DNA Synthesis. As the cell grows larger, the two chromosomes separate. The cell membrane and cell wall then begin to pinch inward, cleaving the cytoplasm. Eventually, the original cell separates into two daughter cells. Each daughter cell receives a copy of the genetic material and can continue growing and dividing.. When nutrients are abundant and conditions are favorable, this process can occur very quickly. Rapid reproduction allows prokaryotic populations to respond quickly to environmental change. Binary fission is a form of asexual reproduction. It produces daughter cells that are genetically identical to the parent cell, except for mutations or newly acquired DNA. Mutations and horizontal gene transfer allow prokaryotes to generate enormous genetic diversity through rapid reproduction, mutation, recombination, and gene transfer.
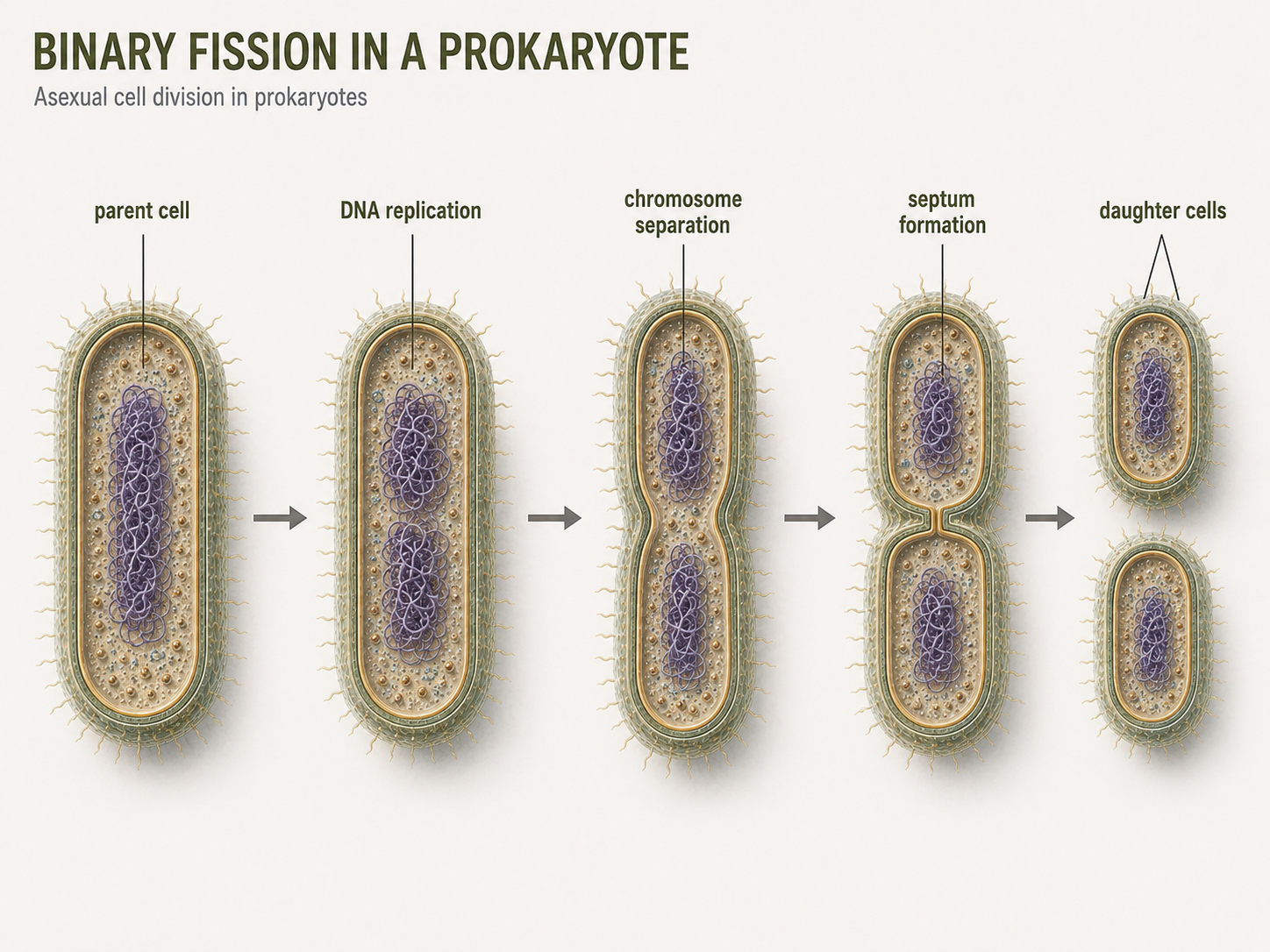
Figure 6. Binary Fission. Many bacteria and archaea reproduce asexually by binary fission, in which one parent cell copies its DNA and divides into two daughter cells. Rapid binary fission allows prokaryotic populations to grow quickly when conditions are favorable.
Cell Envelopes
The cell envelope includes the outer layers of a cell, especially the plasma membrane, cell wall, and any outer structure. Cell envelopes protect cells, help maintain shape, regulate movement of substances, and mediate interactions with the environment. Bacterial cell walls commonly contain peptidoglycan, a molecule made of sugars and short peptides. This is one of the most important structural features of bacterial cells. Bacteria are often described as Gram-positive or Gram-negative based on how their cell envelopes respond to Gram staining. Gram-positive bacteria typically have an inner cell membrane and a thick peptidoglycan layer. Gram-negative bacteria typically have an inner cell membrane, a thinner peptidoglycan layer, and an outer membrane. Archaea cell envelopes are different. Archaea do not contain peptidoglycan in their cell walls. This difference is one of the clearest ways to separate Bacteria from Archaea at the cellular level.
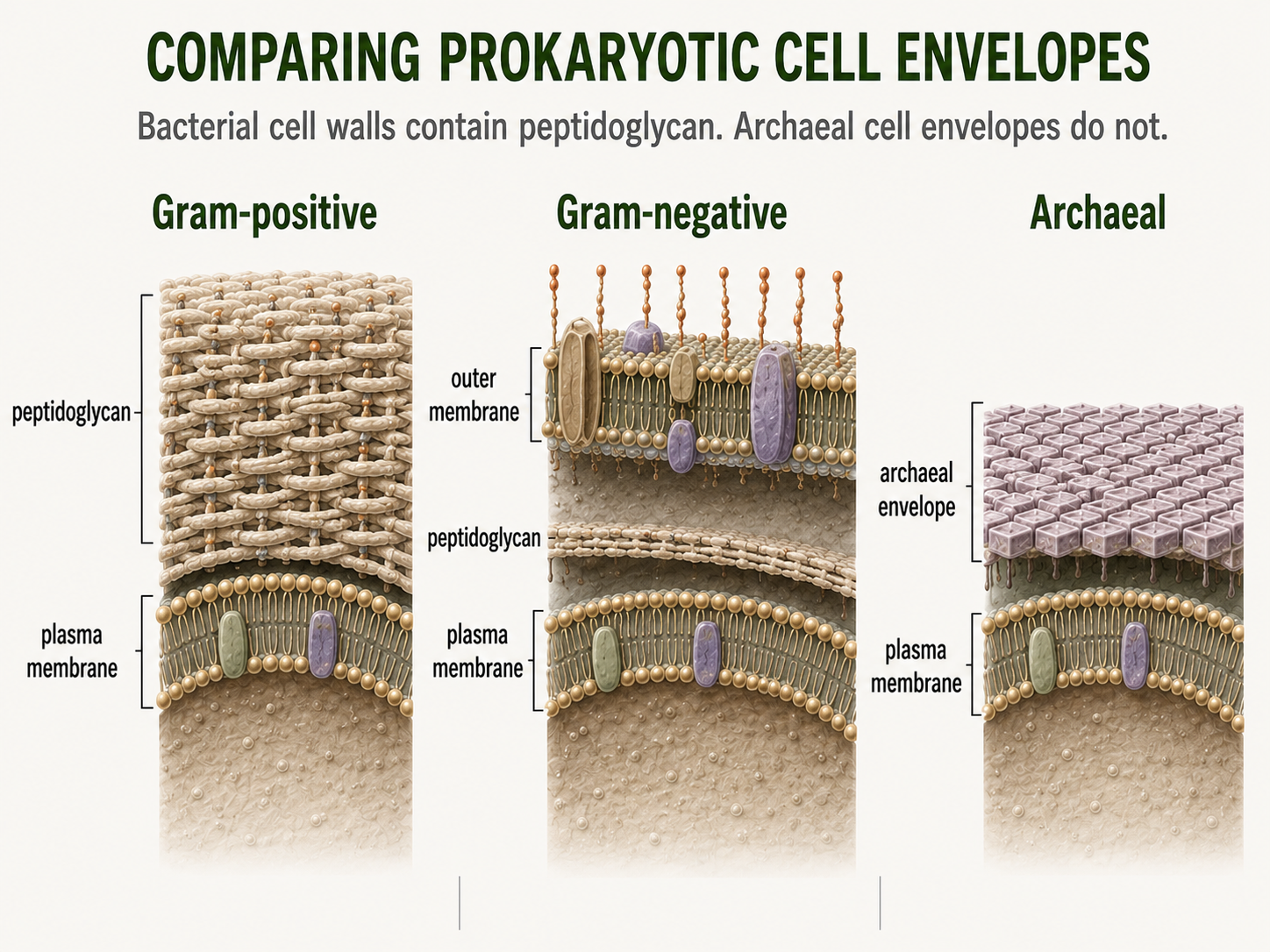
Figure 7. Prokaryotic Cell Envelopes. Cell envelopes protect prokaryotic cells, maintain shape, and help regulate interactions with the environment. Gram-positive bacteria, Gram-negative bacteria, and Archaea differ in envelope structure, especially in the presence or absence of peptidoglycan.
Prokaryotic Movement
Many prokaryotes are not passive cells floating randomly through the world. They can move, attach to surfaces, sense environmental conditions, and respond to changes around them. These abilities help them find nutrients, avoid harmful conditions, colonize new habitats, and interact with other cells. Some bacteria move using flagella, which are long rotating structures that work like propellers. A bacterial flagellum can spin to push or pull the cell through liquid environments. This movement can help bacteria travel toward favorable conditions, such as areas with more nutrients, or away from harmful conditions, such as toxic chemicals. Not all bacterial movement depends on flagella. Some bacteria move by gliding along surfaces, while others use pilus-like structures to pull themselves forward. In this type of movement, a pilus extends outward, attaches to a surface, and then retracts, pulling the cell in that direction. These forms of movement are especially useful on surfaces, including soil particles, rocks, plant roots, host tissues, and biofilms. Archaea also move, but many use structures called archaella. Archaella can look superficially similar to bacterial flagella because both can help cells swim. However, they are structurally and evolutionarily different. Archaella are not simply “archaeal versions” of bacterial flagella. Comparing morphology alone can be misleading in microbes: two structures may look similar and perform similar functions, but have different evolutionary origins. This is true throughout evolutionary history far beyond the prokaryotes.
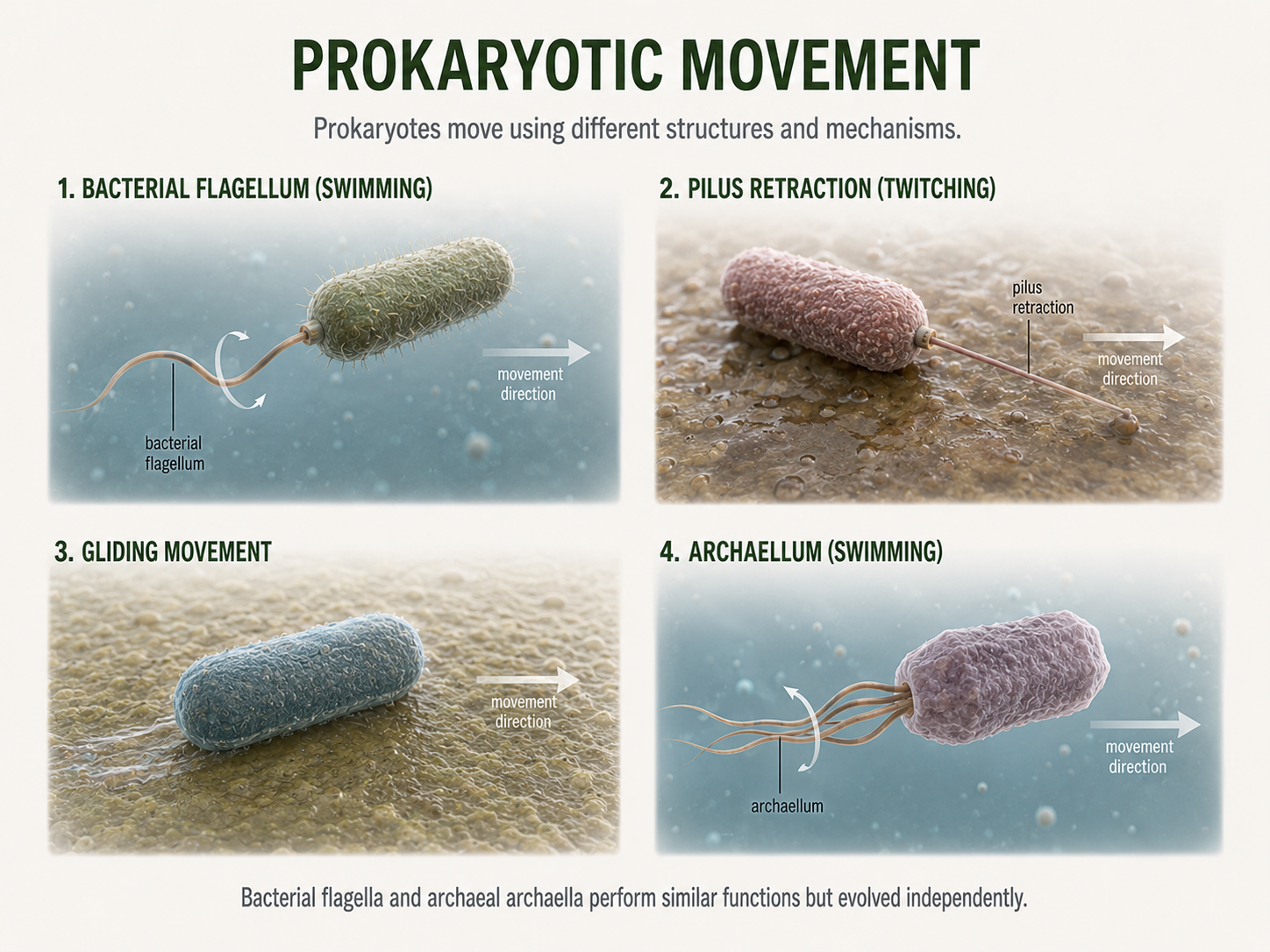
Figure 8. Prokaryotic Movement. Many prokaryotes actively move using structures such as bacterial flagella, retractable pili, gliding mechanisms, or archaeal archaella. Similar-looking movement structures can perform similar functions while having different evolutionary origins.
Prokaryotic Attachments
Many prokaryotes have thin surface structures called fimbriae or pili that extend from the outside of the cell and help them interact with their surroundings. Fimbriae are often short and numerous, attaching cells to surfaces, such as rocks, plant roots, host tissues, or other cells. This attachment can help prokaryotes stay in a favorable environment instead of being washed away. Pili are often longer and less numerous than fimbriae, although the terms are sometimes used differently depending on the organism. Pili can help with attachment, movement, or DNA transfer. Some pili allow cells to pull themselves across surfaces by extending, attaching, and then retracting. Other pili are involved in conjugation, a process in which genetic material moves from one cell to another. During conjugation, a pilus can help bring two cells close enough for DNA to be transferred. These structures show that prokaryotic cell surfaces are active tools for attachment, movement, communication, and gene exchange.
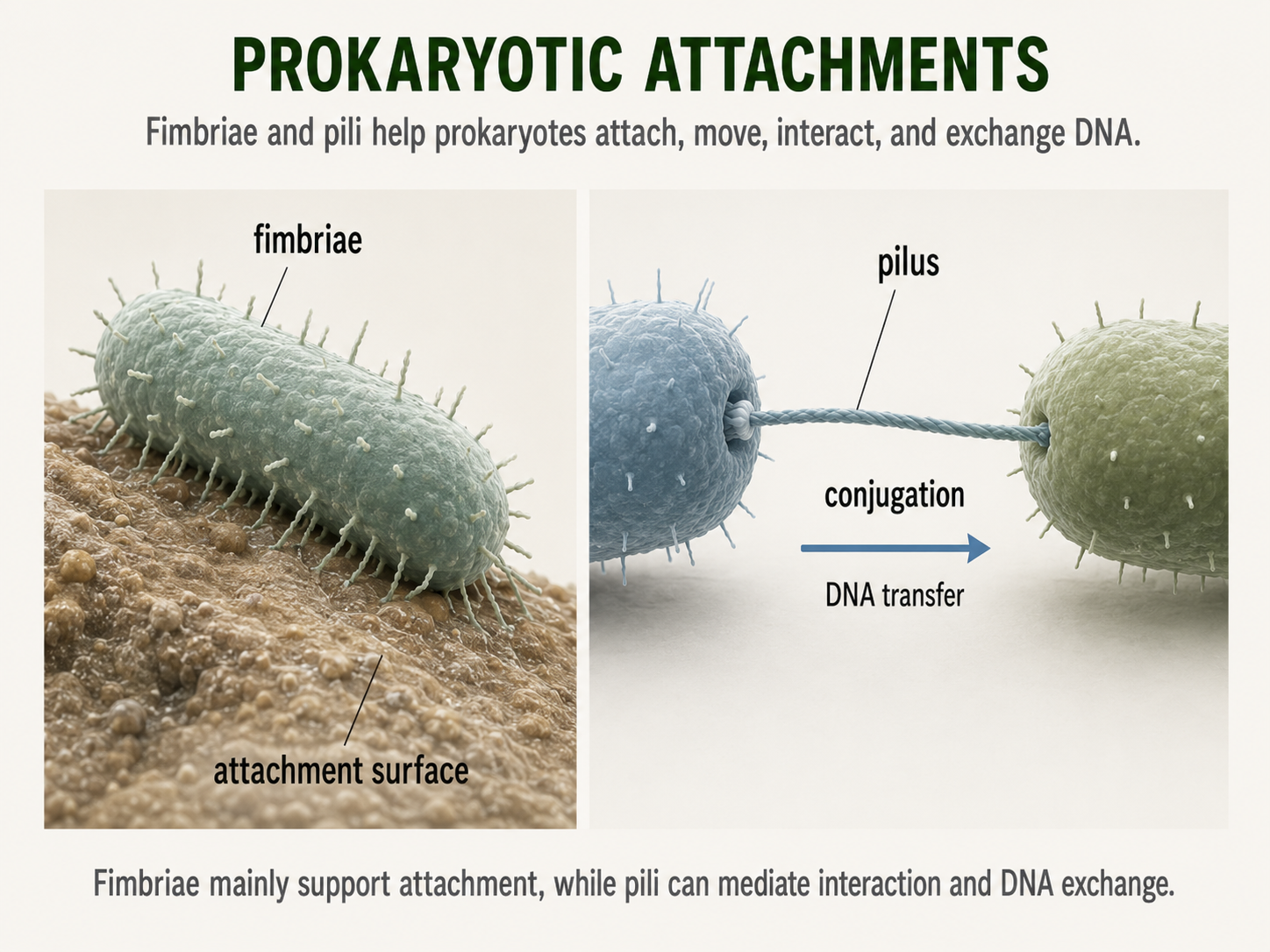
Figure 9. Prokaryotic Attachment Structures. Fimbriae and pili help prokaryotes attach to surfaces, interact with other cells, move across surfaces, and transfer DNA. These external structures make the prokaryotic cell surface an active interface with the environment.
Slime and Biofilms
Some bacteria and archaea produce a sticky outer coating outside the cell wall or cell surface. This coating may be organized as a capsule or may occur as a looser slime layer. These outer layers help cells avoid drying out, attach to surfaces, and form biofilms. In disease-causing bacteria, capsules may also help cells evade immune defenses, making infections harder to clear. A biofilm is a community of microorganisms attached to a surface and embedded in a sticky matrix as a result of intermeshing of the slime layers. Biofilms form on rocks, pipes, teeth, and many other surfaces. Cells within a biofilm may be more resistant to drying, antibiotics, disinfectants, and immune responses. Dental plaque is a familiar, albeit annoying, example of a biofilm. Biofilms show that many bacteria and archaea do not live only as isolated single cells; they often live as interacting microbial communities.
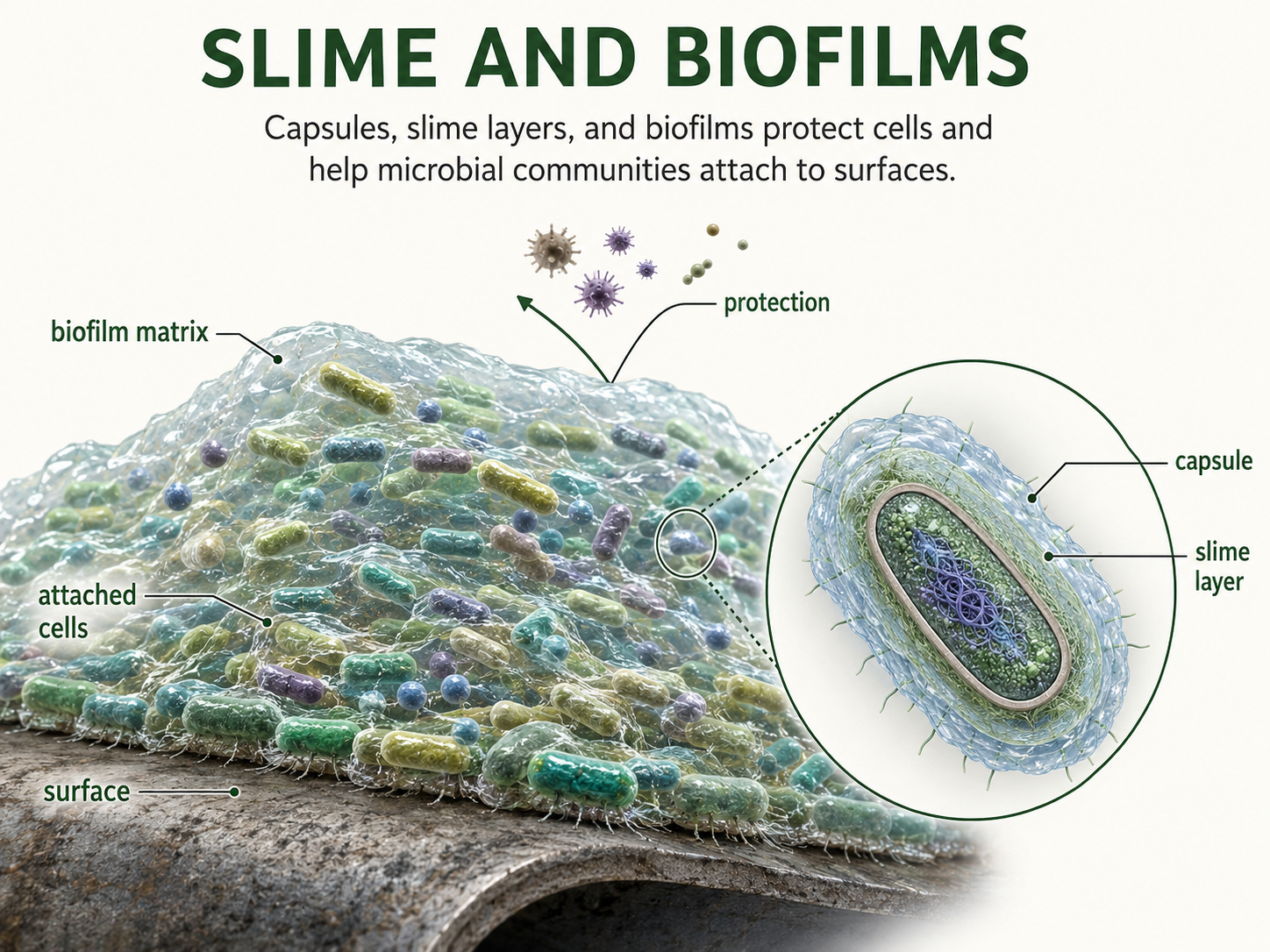
Figure 10. Slime Layers and Biofilms. Some prokaryotes produce capsules or slime layers that help them attach to surfaces, avoid drying out, and form biofilms. Biofilms are microbial communities embedded in a sticky matrix that can protect cells from environmental stress, disinfectants, antibiotics, and immune responses.
From Single Cells to Microbial Communities
Nearly all bacteria and archaea are unicellular, meaning each cell can carry out the basic functions of life on its own. However, this does not mean that prokaryotes always live as isolated individual cells. Many bacteria and archaea are colonial (living in colonies, chains, mats, or biofilms) where cells remain connected and interact with one another. In these communities, cells may share resources, attach to surfaces, communicate chemically, or gain protection from environmental stress. Some bacterial colonies show coordinated behavior and limited cellular specialization. This means that nearby cells may work together or take on slightly different roles, even though they are still prokaryotic cells. But is that multicellularity? Let’s look at an interesting example: photosynthetic bacteria. Cyanobacteria are photosynthetic bacteria that use sunlight to make sugars and release oxygen. Ancient cyanobacteria played a major role in adding oxygen to Earth’s atmosphere, and modern cyanobacteria remain important in aquatic ecosystems and nitrogen cycling. Many cyanobacteria grow as long filaments made of connected cells. In some species, certain cells within the colony become specialized for nitrogen fixation, the process of converting nitrogen gas from the atmosphere into a form that living organisms can use. This division of labor allows some cyanobacterial filaments to coordinate different functions, such as photosynthesis and nitrogen fixation, across connected cells. It could be argued this sharing of resources is a key shift toward what we consider ‘multicellular.’ Let’s look at an even more intriguing example: social bacteria. Myxobacteria are bacteria that can move together in coordinated groups, known as swarm behavior. When conditions are favorable, they hunt other microbes and break down organic material. When conditions become stressful, some myxobacteria gather together and form fruiting-body-like structures. Within these structures, some cells become resistant spores that can survive harsh conditions and later disperse to new environments. This coordinated behavior shows that some bacteria can organize group activity in ways that begin to resemble simple forms of multicellular cooperation. These examples show that prokaryotic life can be highly organized, but it is important not to confuse this with the complex multicellularity seen in animals, plants, and many fungi. Prokaryotes do not form permanent tissues or generate highly integrated body systems. While prokaryotes can be social, colonial and interactive, complex tissue-based multicellularity is mainly a eukaryotic feature.

Figure 11. From Single Cells to Microbial Communities. Although most prokaryotes are unicellular, many live in colonies, chains, mats, filaments, or biofilms. Cyanobacterial filaments and myxobacterial fruiting structures show how prokaryotes can cooperate and display limited division of labor without forming complex tissues.
Prokaryotic Metabolic Diversity
The small size and simple cell organization of prokaryotes can make them seem less complex than eukaryotes. In metabolism, however, bacteria and archaea are remarkably diverse. They can use many different sources of carbon, energy, and electron acceptors, allowing them to survive in environments where many eukaryotes cannot, including oxygen-poor mud, acidic springs, salty lakes, deep-sea vents, and animal intestines. One way to understand prokaryotic metabolism is to separate where cells get their carbon from where they get their energy. An autotroph makes its own organic carbon compounds from carbon dioxide or another inorganic carbon source. A heterotroph obtains organic carbon by consuming or absorbing organic molecules made by other organisms. A phototroph uses light as an energy source, while a chemotroph uses energy stored in chemical compounds. These categories can combine in different ways, which helps explain why prokaryotes are among the most metabolically flexible organisms on Earth. Let’s take a look at some of these unique combinations.
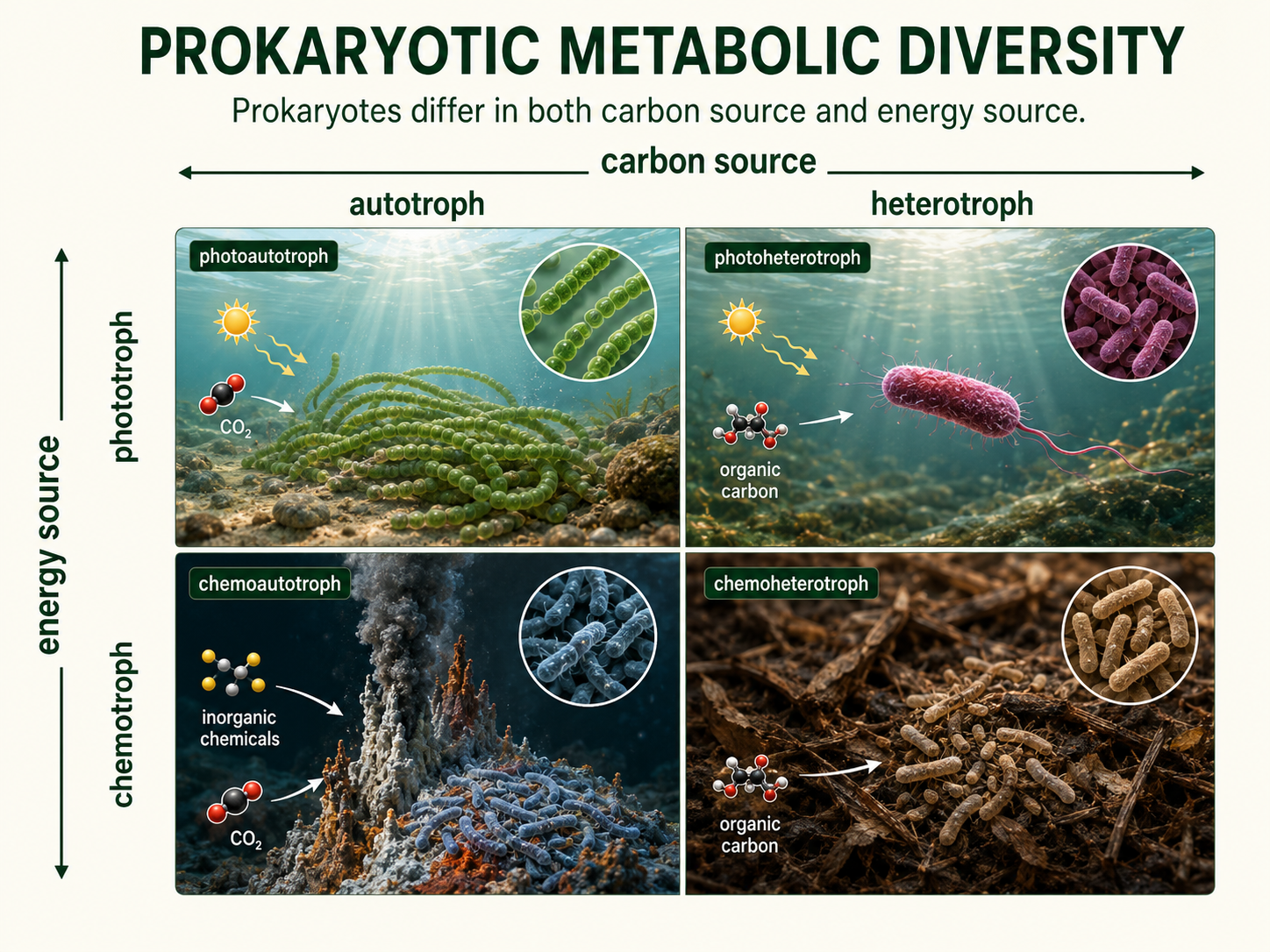
Figure 12. Prokaryotic Metabolic Diversity. Prokaryotes use many combinations of carbon sources and energy sources. Separating carbon source from energy source helps explain why bacteria and archaea can survive in such a wide range of environments.
Metabolic Diversity: Autotrophs
Autotrophs generate their own carbon containing, organic molecules. . Photoautotrophs, such as cyanobacteria, use light energy and carbon dioxide to produce organic molecules. Cyanobacteria are especially important because they perform oxygenic photosynthesis, which releases oxygen gas. Ancient cyanobacteria helped transform Earth’s atmosphere by adding oxygen, making them one of the most important groups of organisms in Earth history. Other prokaryotes are chemoautotrophs, meaning they use chemical energy to build organic molecules from carbon dioxide. An important example of chemoautotrophs are chemolithotrophs, which means they get energy from inorganic substances such as hydrogen sulfide, ammonia, iron, or hydrogen gas. Instead of depending on sunlight, these organisms rely on chemical reactions in their environment. This allows them to live in places such as deep-sea vents, soils, sediments, hot springs, and other habitats where chemical energy supports life. These organisms are especially important in deep-sea vents, soils, sediments, and biogeochemical cycles. For example, some bacteria help convert nitrogen or sulfur compounds from one chemical form to another, making these elements available to other organisms.

Figure 13. Prokaryotic Autotrophs. Some prokaryotes build organic molecules from carbon dioxide using light energy, while others use chemical energy from inorganic substances. Cyanobacteria are photoautotrophs, whereas chemoautotrophs can support ecosystems in places such as vents, sediments, soils, and hot springs.
Metabolic Diversity: Heterotrophs
Many bacteria and archaea are heterotrophs, meaning they obtain carbon and energy by breaking down organic molecules made by other organisms. An important group of heterotrphic prokaryotes are chemoorganotrophs, meaning they get energy and electrons from organic molecules, such as sugars, proteins, fats, dead organisms, or waste products. Many of these organisms are important decomposers because they break down organic matter and recycle nutrients through ecosystems. These prokaryotes are especially important because they act as decomposers, breaking down dead organisms, waste products, fallen leaves, animal remains, and other biological materials. As they decompose this material, they release nutrients back into the environment, where those nutrients can be used again by plants, algae, fungi, microbes, and other organisms. Without this recycling, nutrients would remain locked inside dead material, and ecosystems would eventually lose the raw materials needed for new growth. Decomposition is not just a background process; it is one of the central functions that keeps ecosystems running.

Figure 14. Prokaryotic Heterotrophs. Many bacteria and archaea obtain carbon and energy from organic molecules produced by other organisms. As decomposers, heterotrophic prokaryotes recycle nutrients from dead organisms, waste products, and organic matter back into ecosystems.
Metabolic Diversity: ATP Synthesis
Prokaryotes also vary in how they generate ATP, the usable energy currency of the cell. Some use aerobic respiration, in which oxygen serves as the final electron acceptor at the end of an electron transport chain. This process is efficient and allows cells to produce a relatively large amount of ATP from organic molecules. Other prokaryotes use anaerobic respiration, which also uses an electron transport chain but does not use oxygen as the final electron acceptor. Instead, these organisms may use substances such as nitrate, sulfate, carbon dioxide, or other molecules. This allows them to live in environments where oxygen is absent or limited, such as deep mud, sediments, animal intestines, wetlands, and some aquatic habitats. Still other prokaryotes use fermentation, a process that does not require oxygen or an electron transport chain. Fermentation produces less ATP than respiration, but it allows cells to keep generating energy when suitable electron acceptors are unavailable. Together, aerobic respiration, anaerobic respiration, and fermentation show how flexible prokaryotic metabolism can be.
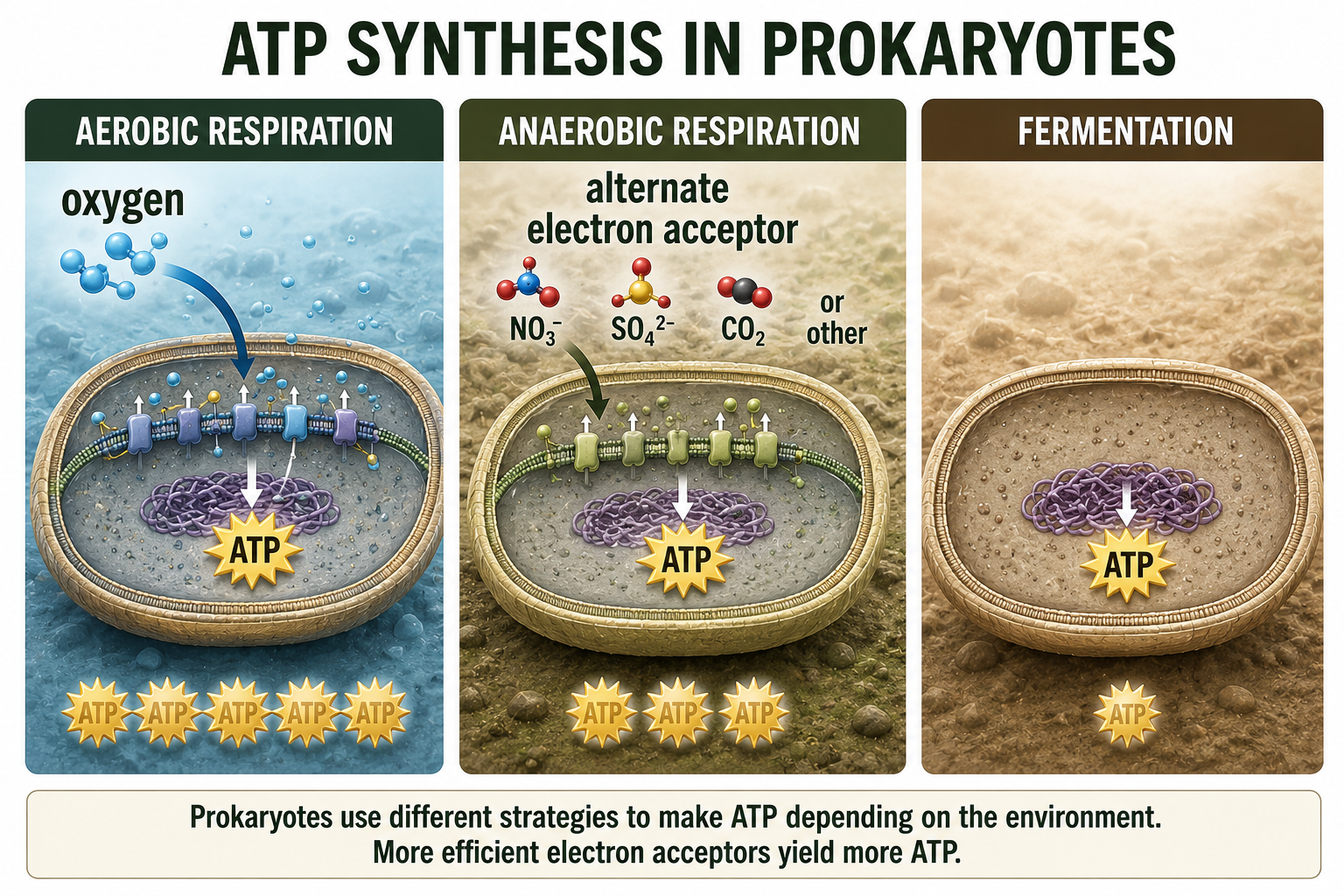
Figure 15. ATP Synthesis in Prokaryotes. Prokaryotes generate ATP through aerobic respiration, anaerobic respiration, or fermentation. These pathways allow different species to live in oxygen-rich, oxygen-poor, or electron-acceptor-limited environments.
Metabolic Diversity: Nitrogen Fixation and Methanogenesis
Bacteria and archaea carry out chemical reactions that are essential to ecosystems. Nitrogen fixation is the process of converting atmospheric nitrogen gas into forms that living organisms can use. Nitrogen gas is abundant in the atmosphere, but most organisms cannot use it directly. This creates a biological problem because nitrogen is needed to build important molecules such as proteins, DNA, and RNA. Certain bacteria and archaea solve this problem by converting nitrogen gas into ammonia or related nitrogen-containing compounds. Once nitrogen has been fixed, it can enter food webs and be used by plants, algae, microbes, and eventually animals. Some nitrogen-fixing prokaryotes live freely in soil or water, while others live in symbiosis with plants. For example, some bacteria live in root nodules of legumes and provide usable nitrogen to the plant in exchange for sugars and shelter. This makes nitrogen-fixing prokaryotes essential to natural ecosystems and agriculture. Methanogenesis is the production of methane by certain archaea called methanogens. Methanogens live in oxygen-free environments, including wetlands, sediments, landfills, rice paddies, and the digestive systems of animals. In these habitats, they help break down organic material under anaerobic conditions, releasing methane as a byproduct. Methanogenesis is important in carbon cycling because it moves carbon through ecosystems in the form of methane. It is also important in climate biology because methane is a powerful greenhouse gas. This process is a useful reminder that Archaea are not simply “unusual bacteria.” Archaea are a separate domain of life with their own evolutionary history, cellular features, and metabolic abilities. Some archaeal metabolisms, including methanogenesis, are distinct from those found in bacteria and eukaryotes. These processes show that prokaryotes may be small, but their chemical activities have major effects on ecosystems, agriculture, and Earth’s atmosphere.

Figure 16. Nitrogen Fixation and Methanogenesis. Bacteria and archaea carry out chemical reactions that shape ecosystems and the atmosphere. Nitrogen-fixing prokaryotes convert nitrogen gas into usable nitrogen compounds, while methanogenic archaea produce methane in oxygen-free environments.
Extremophiles and Ordinary Environments
Because prokaryotes are metabolically flexible, they can live in places that would kill many other organisms. An extremophile is an organism that thrives in extreme conditions, such as extreme heat, high salt, high acidity, high pressure, or lack of oxygen. Some archaea live in hot springs, salt lakes, acidic environments, or deep-sea vents. Some bacteria also thrive in extreme environments. Extremophiles matter because they help scientists understand the limits of life. They also help researchers think about early Earth and the possibility of life beyond Earth. For example, organisms that live without sunlight near hydrothermal vents show that life can be powered by chemical energy rather than solar energy. However, it is misleading to present Archaea as only extremophiles. Many archaea live in ordinary environments, including oceans, soils, sediments, wetlands, and animal digestive systems. Archaea are part of normal microbial communities, including the human microbiome. Current research continues to investigate how human-associated archaea may influence health, but no confirmed archaeal pathogen is known at this time.

Figure 17. Extremophiles and Ordinary Environments. Some prokaryotes thrive in extreme heat, salt, acidity, pressure, or oxygen-free conditions, but bacteria and archaea are also widespread in ordinary habitats. Archaea should not be understood only as extremophiles.
Germ Theory, Sanitation, and Antibiotics
The medical importance of bacteria became much clearer with the development of the germ theory of disease, which states that many diseases are caused by microorganisms. This idea transformed medicine, public health, surgery, food safety, and sanitation. Robert Koch and other scientists developed methods for connecting specific microbes to specific diseases. Koch’s postulates were an early framework for testing whether a microorganism caused a disease, although modern microbiology recognizes exceptions and uses additional molecular and epidemiological tools. Sanitation was one of the most important public health advances in human history. Clean water, sewage treatment, handwashing, food safety, and sterile medical practices dramatically reduced the spread of infectious disease. Antibiotics later added another powerful tool by allowing physicians to treat many bacterial infections directly. Antibiotics are drugs that kill bacteria or inhibit their growth. They do not work against viruses. Antibiotics changed medicine, but their use also created strong selection pressure on bacterial populations. When antibiotics are overused or misused, resistant bacteria are more likely to survive and reproduce. Antibiotic resistance is evolution in action. A bacterial population may already contain a few resistant cells, or resistance may arise by mutation or be acquired through horizontal gene transfer. When an antibiotic kills susceptible cells, resistant cells can survive, reproduce, and pass resistance genes to future generations or sometimes to other bacteria.

Figure 18. Germ Theory, Sanitation, and Antibiotics. Germ theory connected many diseases to microorganisms and transformed medicine and public health. Sanitation reduces microbial transmission, while antibiotics treat bacterial infections but also create selection pressure for antibiotic resistance.
Koch’s Postulates
Koch’s postulates are a set of guidelines used to connect a specific microorganism to a specific disease. They were developed in the late 1800s by Robert Koch and helped establish the germ theory of disease. The basic idea is that if a microbe causes a disease, it should be 1) found in diseased organisms, 2) isolated and grown in culture, 3) cause the same disease when introduced into a healthy host, and then 4) be recovered again from the newly infected host. Koch’s postulates were important because they gave scientists a systematic way to test whether a particular bacterium caused a particular illness. However, they do not apply perfectly to every disease. Some pathogens cannot be grown easily in pure culture, some diseases are caused by multiple organisms, and some people can carry a pathogen without showing symptoms. Even with these limitations, Koch’s postulates remain an important historical and scientific framework for understanding how microbes are linked to disease.
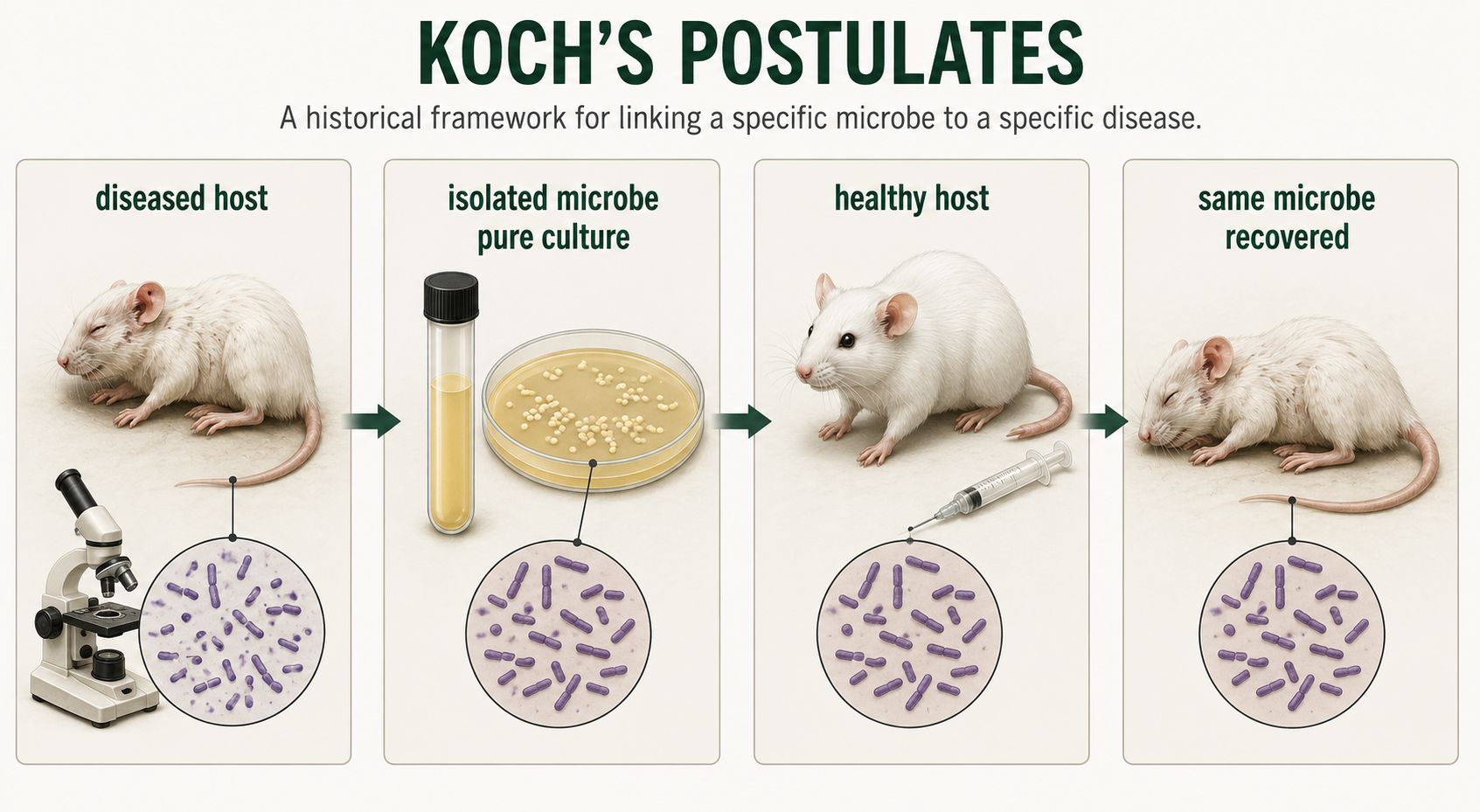
Figure 19. Koch’s Postulates. Koch’s postulates provided an early systematic framework for linking a specific microorganism to a specific disease. Although modern microbiology recognizes many exceptions, the postulates remain historically important for understanding microbial causation.
Discovering the Domains of Life
For much of human history, microbial life was hidden from view. The invention of the microscope made bacteria visible, but it did not make them easy to understand. Under a microscope, many microbes look deceptively simple. They may appear as rods, spheres, spirals, or filaments, but these shapes do not reveal their deep evolutionary relationships. This problem bothered scientists who were trying to classify life. For a long time, all organisms without nuclei were grouped together as “bacteria.” Then, in the 1970s, a scientist named Carl Woese took a different approach. Instead of relying on cell shape or metabolism alone, Woese compared molecules found inside cells. He focused especially on ribosomal RNA, a molecule used by all cells to help build proteins. Because ribosomal RNA is found across life and changes slowly over evolutionary time, it can be used as a molecular record of ancestry. When Woese compared ribosomal RNA sequences from different microbes, he found something surprising. Some prokaryotes that looked like bacteria were not closely related to bacteria at all. They represented a separate major branch of life. These organisms became known as Archaea. Woese’s discovery helped transform biology from the older two-kingdom or five-kingdom view of life into the modern Three Domain System: Bacteria, Archaea, and Eukarya. This discovery changed how scientists study microbes. Today, researchers use 16S rRNA sequencing, genome sequencing, metagenomics, and other molecular tools to identify and compare bacteria and archaea. These methods are especially useful because many microbes cannot be easily grown in a laboratory. Traditional microbiology often depended on culturing organisms, but many bacteria and archaea require specific conditions that are difficult to recreate. Metagenomics allows scientists to study DNA collected directly from environments such as soil, seawater, hot springs, sediments, or the human gut. This approach has revealed enormous microbial diversity that older culture-based methods missed. Prokaryotic classification continues to change as genome data improve. Some bacterial and archaeal group names are revised as scientists discover new relationships. For students, the main lesson is not to memorize every changing name. The important lesson is that molecular evidence transformed microbiology. Carl Woese’s work showed that microbes that look simple under a microscope can have deep and surprising evolutionary histories.
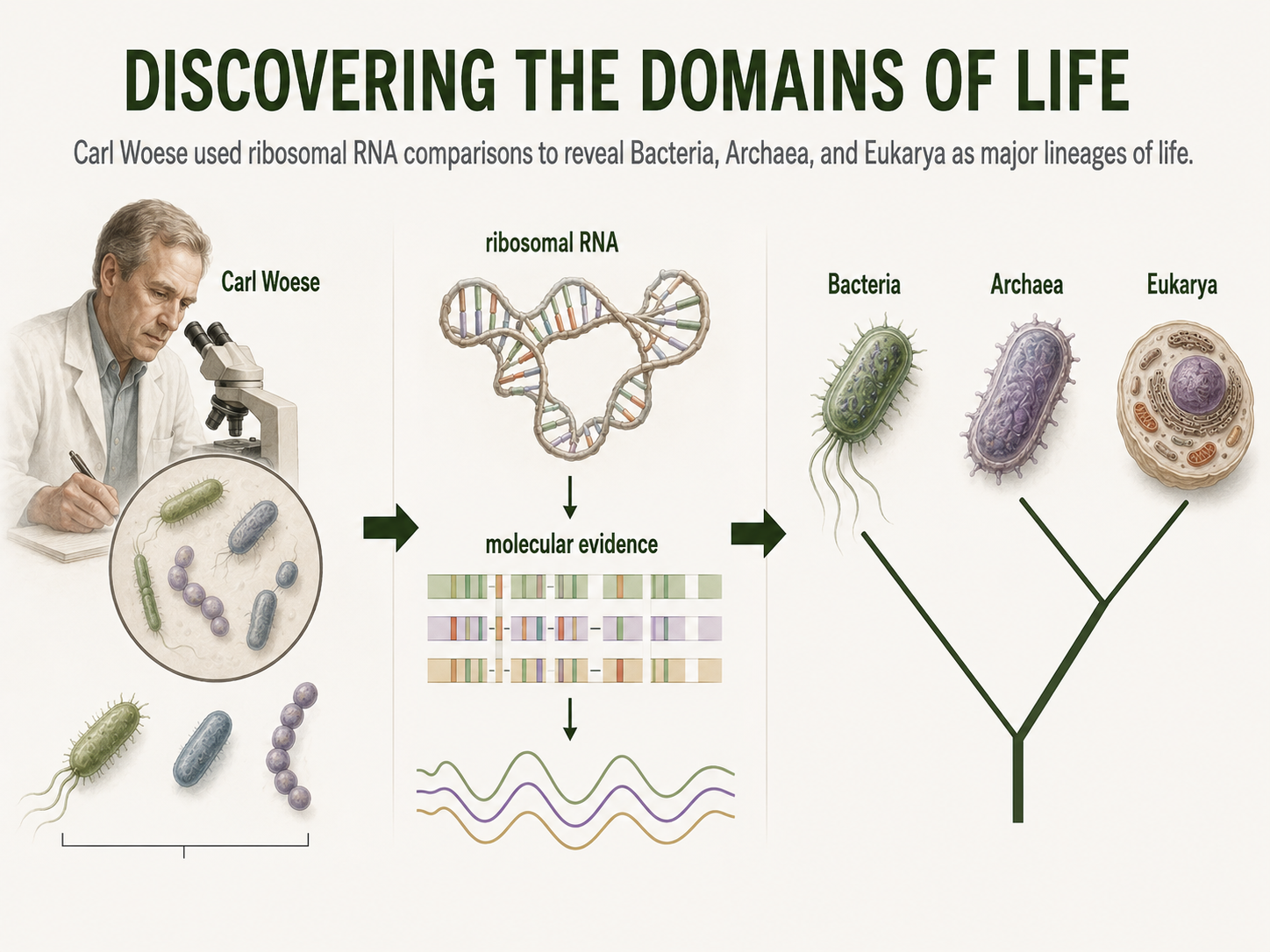
Figure 20. Discovering the Domains of Life. Carl Woese’s comparisons of ribosomal RNA revealed that some prokaryotes once grouped with bacteria belonged to a separate major lineage: Archaea. Molecular evidence transformed biology by showing that microbes with simple shapes can have deep evolutionary differences.
The Three Domains of Life
All living things on Earth share several basic characteristics: a cell membrane, DNA, ribosomes, and proteins. Because these features are found across all life, the earliest common ancestor of modern organisms must have had some version of them. This ancient ancestral cell was probably simple and microscopic, and it may have looked more like a bacterium than a plant, animal, or fungus, although it was not the same as any bacterium alive today. From this early cellular life, natural selection and evolution began shaping organisms in different directions. Over time, populations changed metabolically, genetically, and structurally as they adapted to different environments. One major split produced the lineages that became Bacteria and Archaea. These two groups are both prokaryotic, meaning their cells lack a nucleus, but they differ in important molecular ways, including their cell membrane chemistry, cell wall structure, gene expression, and metabolic pathways. Bacteria became incredibly diverse and adapted to nearly every environment on Earth, while Archaea evolved distinct molecular features and many unusual metabolic abilities, including forms that live in hot springs, salty lakes, oxygen-free sediments, and animal digestive systems. Later in evolutionary history, one branch within the archaeal lineage gave rise to Eukarya. Early eukaryotic cells evolved greater internal complexity, including a nucleus and membrane-bound organelles. A key step was endosymbiosis, when an ancestral archaeal host cell formed a close relationship with a bacterium that eventually became the mitochondrion. This evolutionary partnership helped produce the eukaryotic lineage, which later diversified into protists, fungi, plants, and animals.
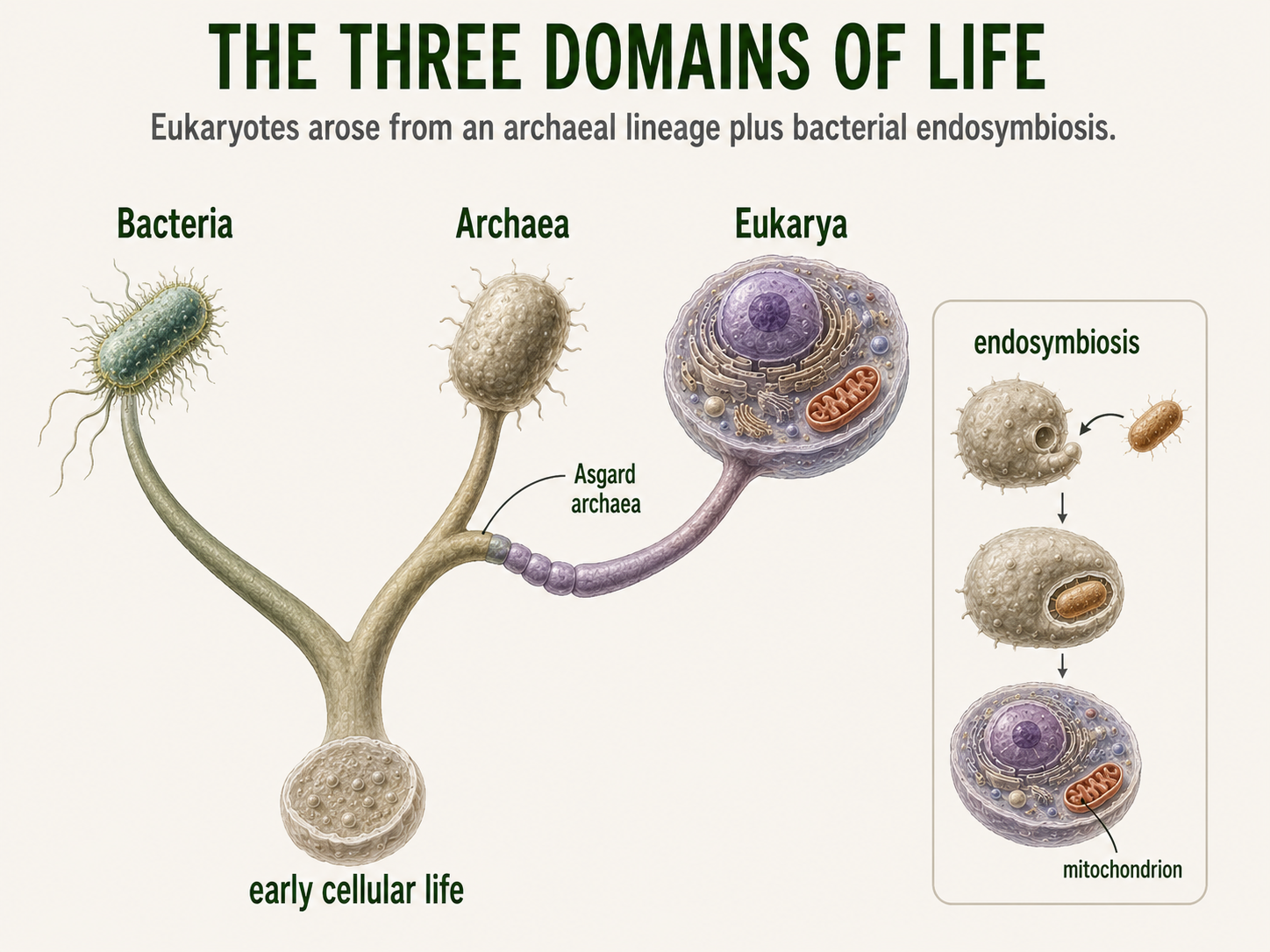
Figure 21. The Three Domains of Life. Bacteria, Archaea, and Eukarya share ancient cellular features but differ in major molecular and structural traits. Modern evidence suggests that Eukarya arose from an archaeal lineage and gained mitochondria through bacterial endosymbiosis.
Bacteria Diversity
Domain: Bacteria
After the earliest split among the major domains of life, the bacterial lineage continued to branch into many deeply divergent groups. The next major branching pattern within Domain Bacteria is not as simple or universally agreed upon as the division between Bacteria, Archaea, and Eukarya, but scientists recognize several major bacterial lineages that represent deep evolutionary diversity. As bacteria spread across ancient Earth, different lineages began solving different survival problems.
One major branch, often called Terrabacteria, became especially successful in places where cells had to survive sunlight, oxygen, drying, and life on land or in soil. Some members of this lineage, the cyanobacteria, changed the planet by evolving oxygen-producing photosynthesis. As they used sunlight to make food, they released oxygen into the atmosphere and helped reshape Earth’s environments. Other Terrabacteria became masters of the soil. Actinomycetes formed branching networks through the ground and produced many of the antibiotics that humans later discovered and used in medicine. Firmicutes followed another path, with some species evolving tough spores that allow them to survive harsh conditions, while others became important residents of animal digestive systems.
A different broad branch, often called Gracilicutes, included bacteria with thinner cell walls and outer membranes. This lineage produced groups such as Proteobacteria, one of the most diverse bacterial groups on Earth. Some Proteobacteria fix nitrogen, some use sulfur compounds for energy, some live in water or soil, and others, such as Escherichia coli, became familiar inhabitants of animal intestines. Bacteroidetes also became widespread, especially in soils, oceans, and digestive systems, where many species help break down complex organic material. Other bacterial lineages followed even more unusual evolutionary paths. Spirochetes evolved long spiral-shaped bodies that move with a corkscrew motion. Chlamydiae became specialists at living inside host cells. Planctomycetes developed unusual internal cell organization. Deinococcus-Thermus includes bacteria able to survive extreme stress, including radiation, heat, and drying.
Together, these lineages show that bacterial diversity is not mainly a story of visible body forms. It is a story of ancient cells evolving different ways to survive, feed, move, reproduce, and transform the planet. This diversity shows that bacteria are not a single simple type of organism. They are an ancient, deeply branching domain whose members evolved many different metabolisms, cell structures, and ecological roles. Some bacteria became photosynthetic, some became decomposers, some became symbionts, some became pathogens, and many became essential partners in ecosystems. The bacterial tree of life therefore reveals that most bacterial diversity is hidden at the molecular and metabolic level rather than in obvious body form.
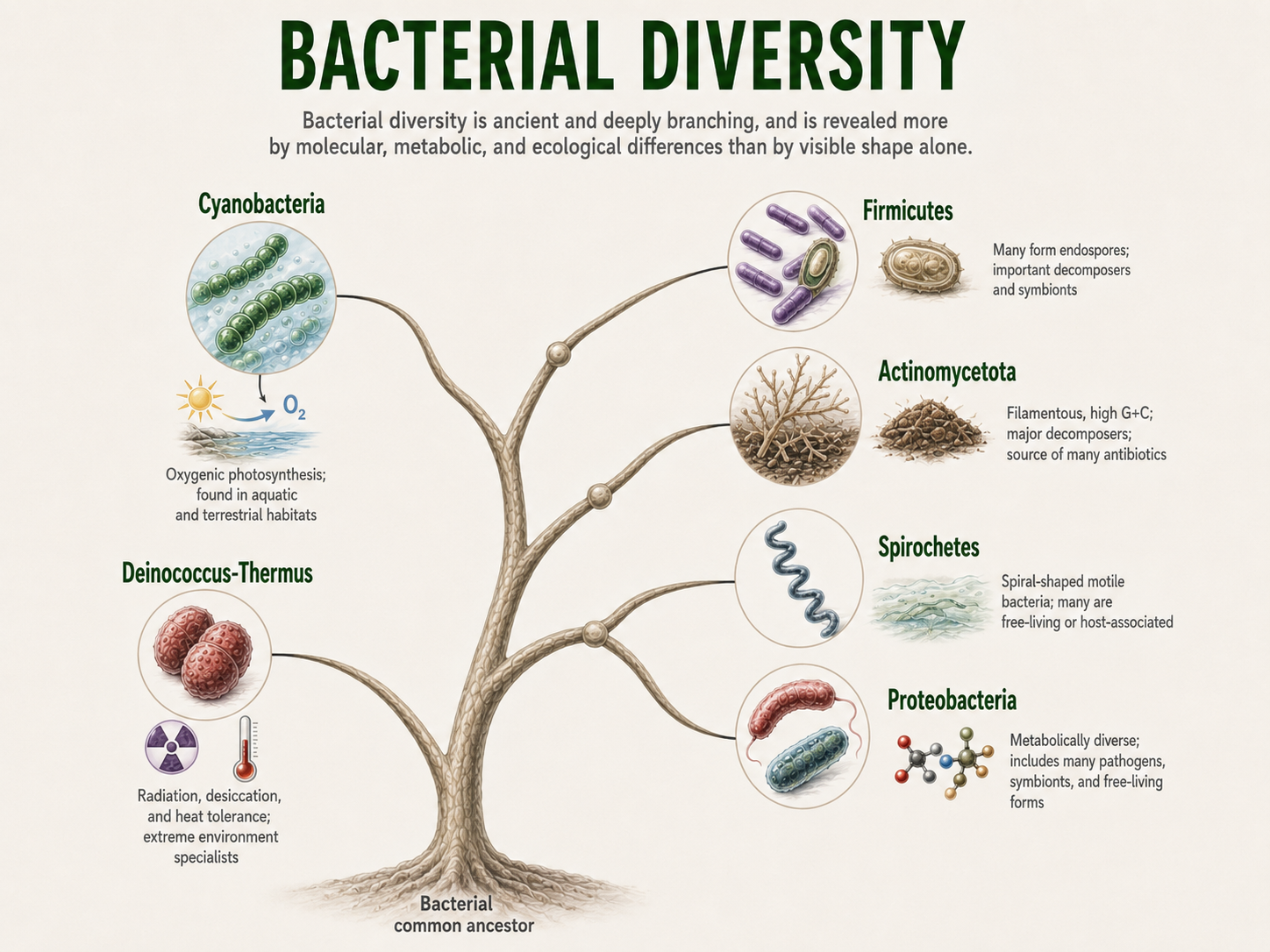
Figure 22. Bacterial Diversity. Bacteria are an ancient and deeply branching domain whose diversity is expressed through metabolism, ecology, cell structure, and evolutionary history. Major lineages include cyanobacteria, actinomycetes, firmicutes, proteobacteria, spirochetes, and other groups with distinct ecological roles.
Archaea Diversity
Domain: Archaea
Archaea are metabolically diverse, molecularly distinct, ecologically important, and central to the story of how complex cells evolved. After the earliest split among the major domains of life, the archaeal lineage also began branching into many deeply different groups. Archaea are often less familiar than bacteria because they are not commonly associated with human disease, and many are difficult to grow in the laboratory. For a long time, they were thought of mostly as organisms from extreme environments, but modern genome studies have shown that archaea are widespread in ordinary habitats as well. Like bacteria, archaeal diversity is not mainly a story of visible body shapes. It is a story of ancient cells evolving unusual membranes, distinct molecular machinery, and many different ways of living. Some archaeal lineages became specialists in oxygen-free environments. Methanogens evolved a metabolism that produces methane, allowing them to survive in places where oxygen is absent. They are common in wetlands, sediments, landfills, and the digestive systems of animals, where they help break down organic material and release methane as a byproduct. Other archaea followed a different path and became adapted to salty environments. Halophiles thrive in places such as salt lakes, salt flats, and highly saline ponds, where most cells would lose water and struggle to survive. These organisms show how archaeal membranes and proteins can be shaped by natural selection to function under conditions that would be stressful or deadly for many other forms of life. Other archaeal groups became masters of heat. Thermophiles live in hot springs, geothermal soils, and hydrothermal vents, where temperatures can be high enough to damage ordinary proteins and membranes. Their cells help scientists understand how life can remain stable under extreme conditions and where the limits of life may be. Archaeans are easy to overlook because they are microscopic, but their metabolic activity affects entire ecosystems.

Figure 23. Archaea Diversity. Archaea are a distinct domain of life with unique molecular features, unusual membrane chemistry, and diverse metabolisms. Although some live in extreme environments, many archaea are common in oceans, soils, sediments, wetlands, and animal digestive systems.
Archaean Ancestors and Eukaryotics Roots
Domains: Archaea and Eukarya
One of the most important archaeal lineages for understanding the history of life is the Asgard archaea. These archaea are especially interesting because molecular and genome-based studies place them close to the lineage that gave rise to eukaryotes, the group that includes protists, fungi, plants, and animals. This discovery changed how scientists think about the origin of complex cells. For many years, eukaryotes were treated as a completely separate kind of life, distinct from both bacteria and archaea. Modern evidence suggests a more connected story: eukaryotic cells likely emerged from within, or very close to, an ancient archaeal lineage. This does not mean that modern Asgard archaea are “almost eukaryotes” or halfway between archaea and animals. They are modern organisms with their own long evolutionary history. However, their genomes contain genes that resemble some of the genes eukaryotic cells use to control internal structure, membrane shape, and cellular organization. These genes provide clues about the ancient archaeal host cell that eventually became part of the eukaryotic story. In other words, Asgard archaea help scientists reconstruct what the archaeal ancestor of eukaryotes may have been like. The origin of eukaryotic cells also involved a major evolutionary partnership called endosymbiosis. At some point, an archaeal host cell formed a close relationship with a bacterium. Instead of being digested or destroyed, that bacterium remained inside the host and eventually evolved into the mitochondrion. Mitochondria became the energy-producing organelles found in nearly all eukaryotic cells today. This partnership was one of the most important events in the history of life because it helped make larger, more complex cells possible. The roots of eukaryotic life reach back into Archaea, while one of the most important engines of eukaryotic complexity came from Bacteria.

Figure 24. Archaean Ancestors and Eukaryotic Roots. Asgard archaea help scientists reconstruct the archaeal ancestry of eukaryotic cells. Eukaryotic origins involved an archaeal host lineage and a bacterial endosymbiont that eventually evolved into the mitochondrion.