
Chapter: Plants
Characteristics of Plants
Domain Eukarya: Archaeplastida: Viridiplantae: Embryophytes
Plants are eukaryotic organisms whose evolutionary history is rooted in photosynthesis, multicellularity, and life on land. Most plants are photoautotrophs, meaning they use light energy to build sugars from carbon dioxide and water. Their cells usually contain chloroplasts, organelles that carry out photosynthesis, and most plant cells are surrounded by cellulose-rich cell walls that provide support and protection. In this chapter, the word “plant” will mainly refer to land plants, also called embryophytes. Land plants include liverworts, mosses, hornworts, ferns and their relatives, gymnosperms, and flowering plants. These groups share several important traits. They have chloroplasts derived from an ancient cyanobacterial endosymbiont, cell walls made largely of cellulose, multicellular bodies, and a life cycle that alternates between a haploid gametophyte generation and a diploid sporophyte generation. Land plants are also called embryophytes because the young embryo is retained and nourished by parental tissue, a major reproductive innovation that helped distinguish land plants from their algal relatives. Plants are not simply “green organisms.” They are an evolutionary lineage shaped by the challenges of life in sunlight, water, air, and soil. The history of plants is the story of how photosynthetic organisms moved from aquatic environments onto land and evolved structures that allowed them to resist drying, exchange gases, move water internally, grow upright, reproduce without standing water, and form complex relationships with animals, fungi, and ecosystems.
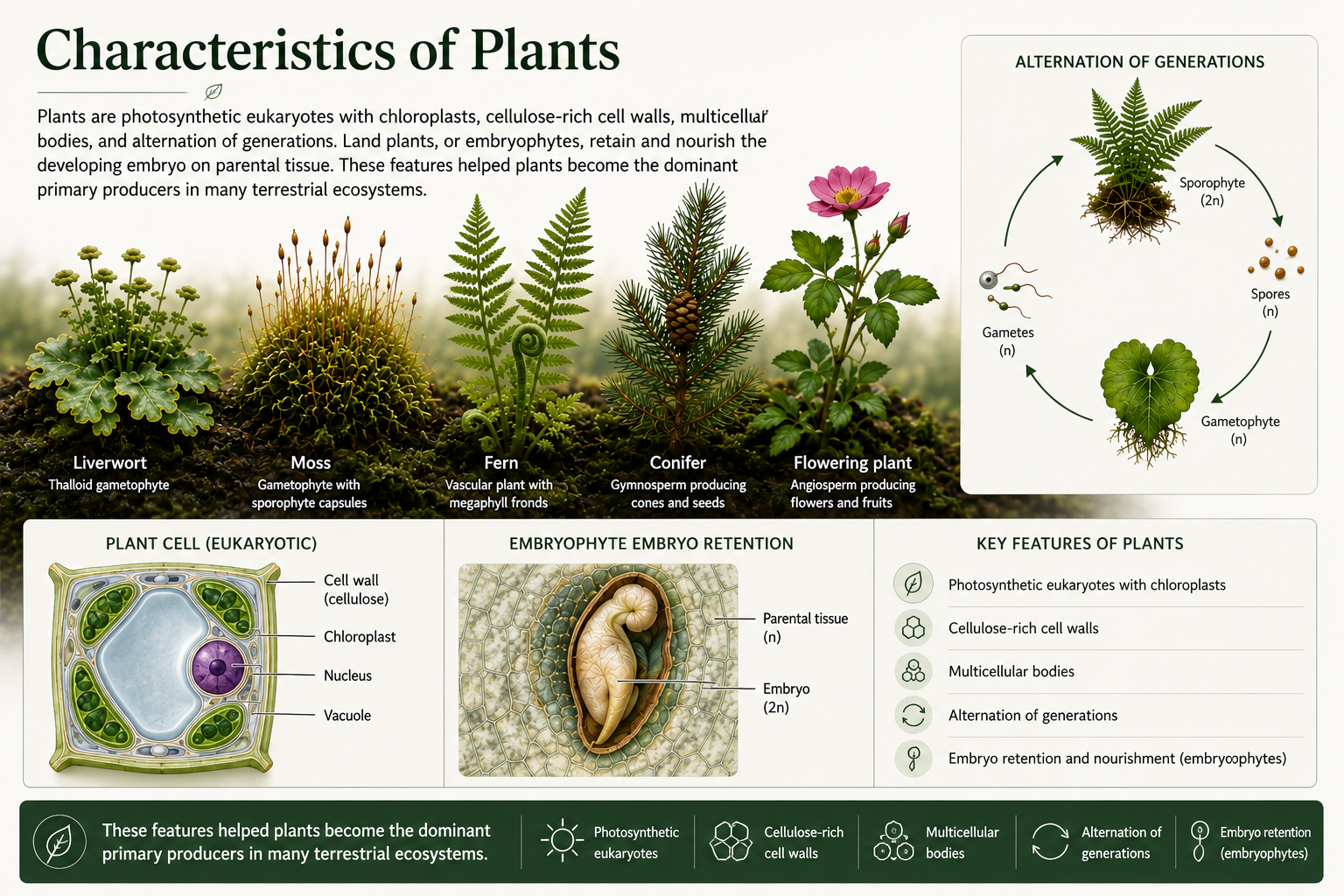
Figure 1. Characteristics of Plants. Plants are photosynthetic eukaryotes with chloroplasts, cellulose-rich cell walls, multicellular bodies, and alternation of generations. Land plants, or embryophytes, retain and nourish the developing embryo on parental tissue. These features helped plants become the dominant primary producers in many terrestrial ecosystems.
Archaeplastida: The Origin of Photosynthetic Eukaryotes
Domain Eukarya: Clade Archaeplastida
Plants belong within Archaeplastida, a major eukaryotic lineage that also includes glaucophyte algae, red algae, green algae, and land plants. The defining evolutionary event in this lineage was the origin of the chloroplasts. Chloroplasts originated when an early eukaryotic cell engulfed a photosynthetic cyanobacterium and retained it instead of digesting it. Over time, the cyanobacterium became an internal organelle that supplied sugars to the host cell through photosynthesis. This event is called primary endosymbiosis because the eukaryotic host engulfed a cyanobacterium directly. This is different from secondary endosymbiosis, which occurs when one eukaryote engulfs another photosynthetic eukaryote. Primary endosymbiosis gave rise to the plastids of Archaeplastida, including the chloroplasts of red algae, green algae, and land plants. Modern chloroplasts still show signs of this origin, including their double membranes and genetic similarities to cyanobacteria.
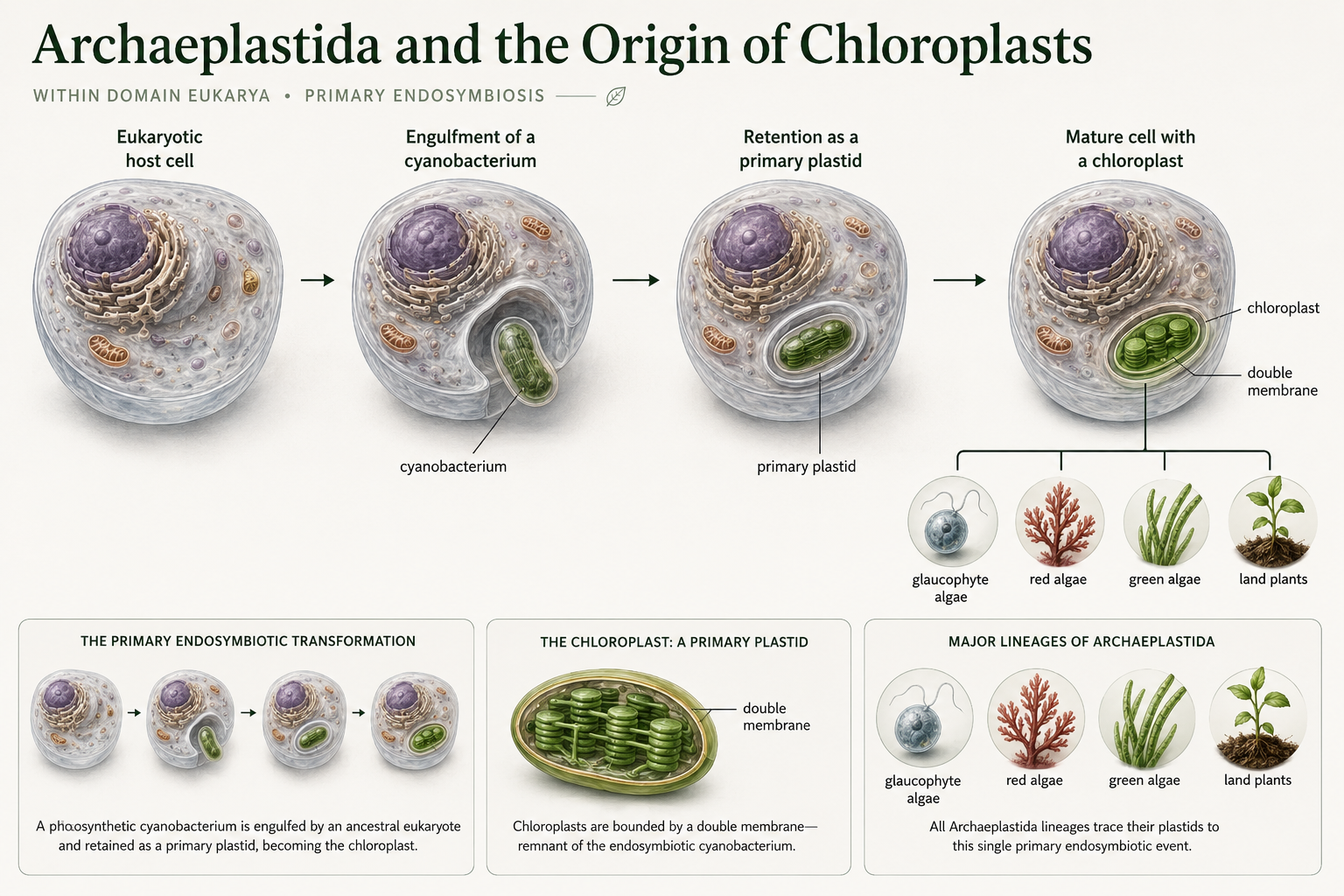
Figure 2. Archaeplastida and the Origin of Chloroplasts. Archaeplastida includes glaucophyte algae, red algae, green algae, and land plants. This lineage originated after a eukaryotic ancestor engulfed a cyanobacterium through primary endosymbiosis. The retained cyanobacterium became the plastid lineage that includes modern chloroplasts.
Glaucophyte Algae: A Window into Early Plastid Evolution
Domain Eukarya: Archaeplastida: Phylum Glaucophyta
Glaucophyte algae (Phylum Glaucophyta) are a small group of mostly freshwater, single-celled algae. They are important because they retain features that help biologists understand early plastid evolution. Their plastids are sometimes called cyanelles because they still resemble cyanobacteria in some ways more strongly than the chloroplasts of green plants do. Glaucophytes photosynthesize using chlorophyll a and accessory pigments called phycobiliproteins. Unlike land plants and green algae, they do not have chlorophyll b as a major photosynthetic pigment. This makes them useful for comparing the major archaeplastid lineages. Glaucophytes are not “primitive plants” in the sense of being unchanged ancestors, but they preserve traits that help reveal what early photosynthetic eukaryotes may have been like.
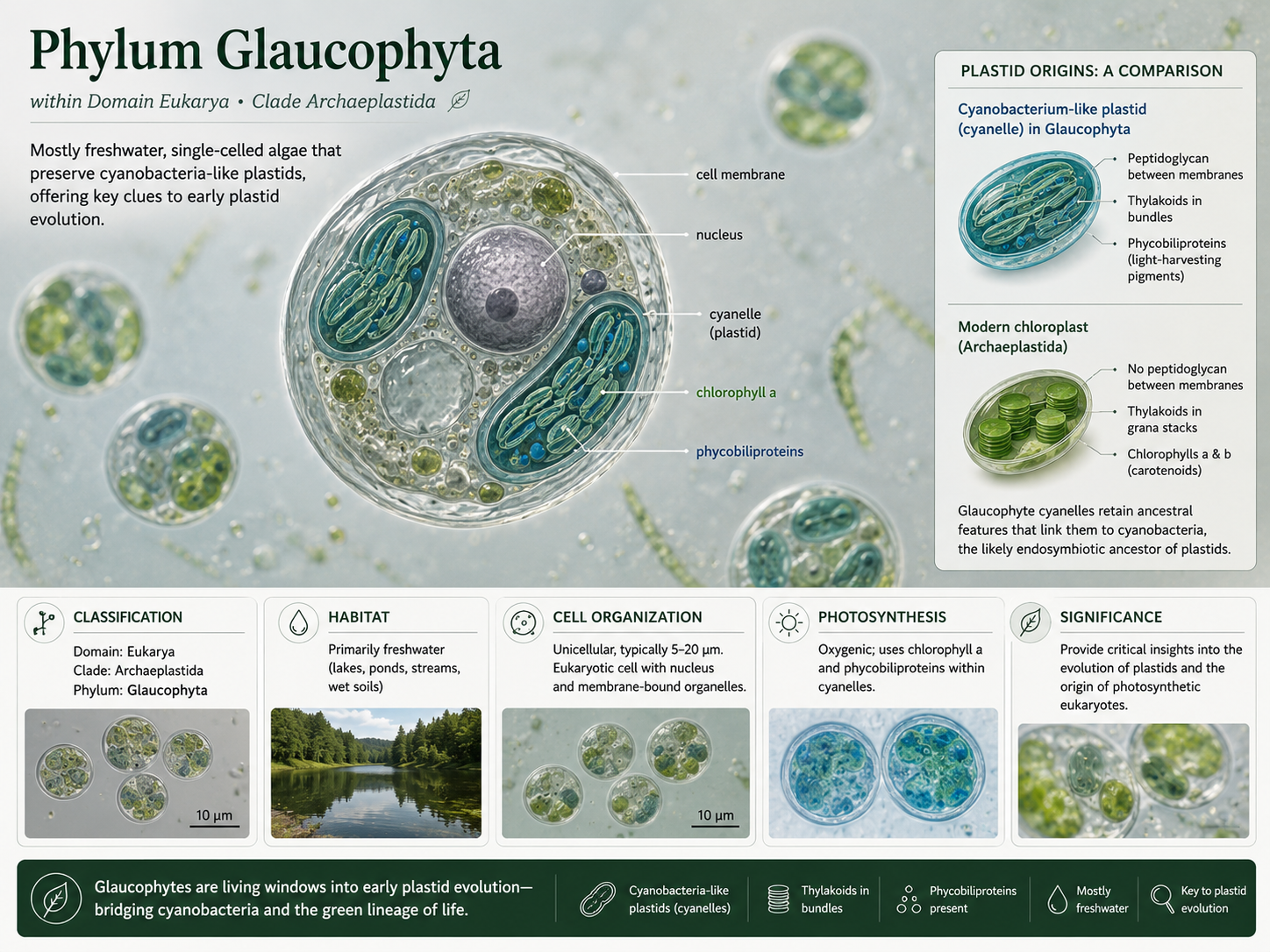
Figure 3. Phylum Glaucophyta. Glaucophyte algae are single-celled freshwater archaeplastids with plastids that retain cyanobacteria-like features. Their pigment system includes chlorophyll a and phycobiliproteins, making them useful for understanding early plastid evolution.
Red Algae
Domain Eukarya: Archaeplastida: Phylum Rhodophyta
Red algae belong to Phylum Rhodophyta and are one of the major lineages within Archaeplastida. Most red algae are marine, and many are multicellular. They are especially abundant in coastal and reef environments, where they contribute to primary production and provide habitat for other organisms. Red algae use chlorophyll a and phycobiliproteins for photosynthesis. These pigments allow many red algae to absorb wavelengths of light that penetrate deeper into water than the light most useful to green plants. This helps explain why red algae are often successful in marine environments, including deeper or shaded habitats. Many red algae appear red, pink, or purple because their accessory pigments reflect reddish wavelengths. Red algae are also important in the fossil record and in human life. Some ancient red algae provide evidence of early multicellular eukaryotes. Living red algae include forms used in food, agar, carrageenan, and marine habitat formation. Like glaucophytes, red algae are not land plants, but they are part of the broader photosynthetic lineage that helps explain the origin and diversification of plant-like life.

Figure 4. Phylum Rhodophyta. Red algae are mostly marine archaeplastids that use chlorophyll a and phycobiliproteins. Their accessory pigments help them photosynthesize in aquatic environments where light conditions differ from those on land. Many red algae are multicellular and ecologically important in coastal ecosystems.
Viridiplantae: Green Algae and Land Plants
Domain Eukarya: Archaeplastida: Viridiplantae
Green algae and land plants together belong to Viridiplantae, the green plant lineage. This group is defined by chloroplasts that contain chlorophyll a and chlorophyll b, along with cell walls that usually contain cellulose. Chlorophyll a and chlorophyll b absorb red and blue wavelengths of light more strongly than green wavelengths, which is why many green algae and land plants appear green. The term “green algae” is useful, but it can also be misleading. Green algae do not form a single simple ladder leading directly to land plants. Instead, the green plant lineage split into major branches. One branch includes many green algae traditionally placed in Phylum Chlorophyta. Another branch, the Streptophyta, includes several groups of freshwater green algae and the land plants. Land plants evolved from within the streptophyte side of the green plant lineage, not from modern chlorophyte algae. This distinction matters because students often imagine evolution as one modern group turning into another modern group. That is not how evolution works. Modern green algae are not ancestors of modern plants. Instead, green algae and land plants share common ancestors, and some living streptophyte algae retain traits that help us infer what the algal relatives of early land plants were like.

Figure 5. Viridiplantae. Green algae and land plants belong to Viridiplantae, the green plant lineage. Members of this group typically have chlorophyll a, chlorophyll b, and cellulose-rich cell walls. Land plants evolved from within the streptophyte branch of green plants rather than from modern chlorophyte algae.
Streptophyte Algae: Close Relatives of Land Plants
Domain Eukarya: Archaeplastida: Viridiplantae: Streptophyta
Streptophyta includes land plants and their closest algal relatives. These algal relatives are often called streptophyte algae or charophyte algae. They are mostly freshwater organisms, and they are important because they share several cellular and reproductive traits with land plants. Stoneworts, placed in Class Charophyceae and Order Charales in many classifications, are one familiar group of streptophyte algae. They are freshwater, multicellular algae with branching bodies that can superficially resemble small aquatic plants. Their bodies may have stem-like axes and whorls of branchlets around nodes. They also have multicellular reproductive structures, including sperm-producing antheridia and egg-containing oogonia. Older teaching treatments often described stoneworts as the closest living relatives of land plants. More recent molecular studies have often supported Zygnematophyceae, a different group of streptophyte algae that includes forms such as Spirogyra and desmids, as the closest living sister lineage to land plants. Land plants evolved from within the streptophyte green plant lineage. Stoneworts remain useful examples to visualize the transition to land, because their body organization and reproductive structures help to similarities between freshwater green algae and land plants.

Figure 6. Streptophyte Algae and the Origin of Land Plants. Streptophyte algae include several freshwater green algal lineages closely related to land plants. Stoneworts are useful teaching examples because they have plant-like branching bodies and multicellular reproductive structures, although modern molecular evidence often places Zygnematophyceae as the closest living sister lineage to land plants.
Origin of Land Plants: The Embryophytes
Domain Eukarya: Archaeplastida: Viridiplantae: Clade Embryophyta
Land plants are embryophytes, meaning they protect and nourish the developing embryo on the parent plant. This was one of the most important transitions in plant evolution. In aquatic algae, reproduction often occurs directly in water, where cells and embryos are protected from drying. On land, reproductive cells and young embryos face a much harsher environment. They can dry out, be exposed to ultraviolet radiation, and lose direct access to water. Early land plants evolved several adaptations that helped solve these problems. A waxy cuticle reduced water loss from exposed surfaces. Stomata, small adjustable openings in the plant surface, allowed gas exchange while helping regulate water loss. Spores with tough cell walls resisted dehydration. Multicellular sporangia protected developing spores. Multicellular gametangia protected gametes. Most importantly, the embryo was retained on parental tissue, where it received protection and nutrients during early development. These features did not make early land plants fully independent of water. Many early-diverging land plants still require moisture for reproduction because their sperm must swim to the egg. But the origin of the embryophyte body plan allowed photosynthetic organisms to begin colonizing terrestrial environments in a new way.

Figure 7. Origin of Land Plants. Land plants evolved from streptophyte algal ancestors and became adapted to terrestrial life through features such as a cuticle, stomata, protected reproductive structures, resistant spores, and embryos retained on parental tissue.
Alternation of Generations
Domain Eukarya: Archaeplastida: Viridiplantae: Embryophyta
All land plants have a life cycle called alternation of generations. This means that the life cycle alternates between two multicellular stages: a haploid gametophyte and a diploid sporophyte. Haploid cells have one set of chromosomes. Diploid cells have two sets of chromosomes. In plants, the gametophyte produces gametes via mitosis, not meiosis. When sperm and egg fuse during fertilization, they form a diploid zygote. The zygote develops into the sporophyte. The sporophyte produces haploid spores by meiosis. These spores grow into new gametophytes. This pattern is different from the animal life cycle emphasized in the animal chapter. In animals, the multicellular body is usually diploid, and the haploid phase is reduced to sperm and eggs. In plants, both the haploid and diploid phases can be multicellular, although their relative size and independence vary across plant groups. In liverworts, mosses, and hornworts, the gametophyte is the dominant, most visible stage. In vascular plants, including ferns, gymnosperms, and flowering plants, the sporophyte becomes dominant.
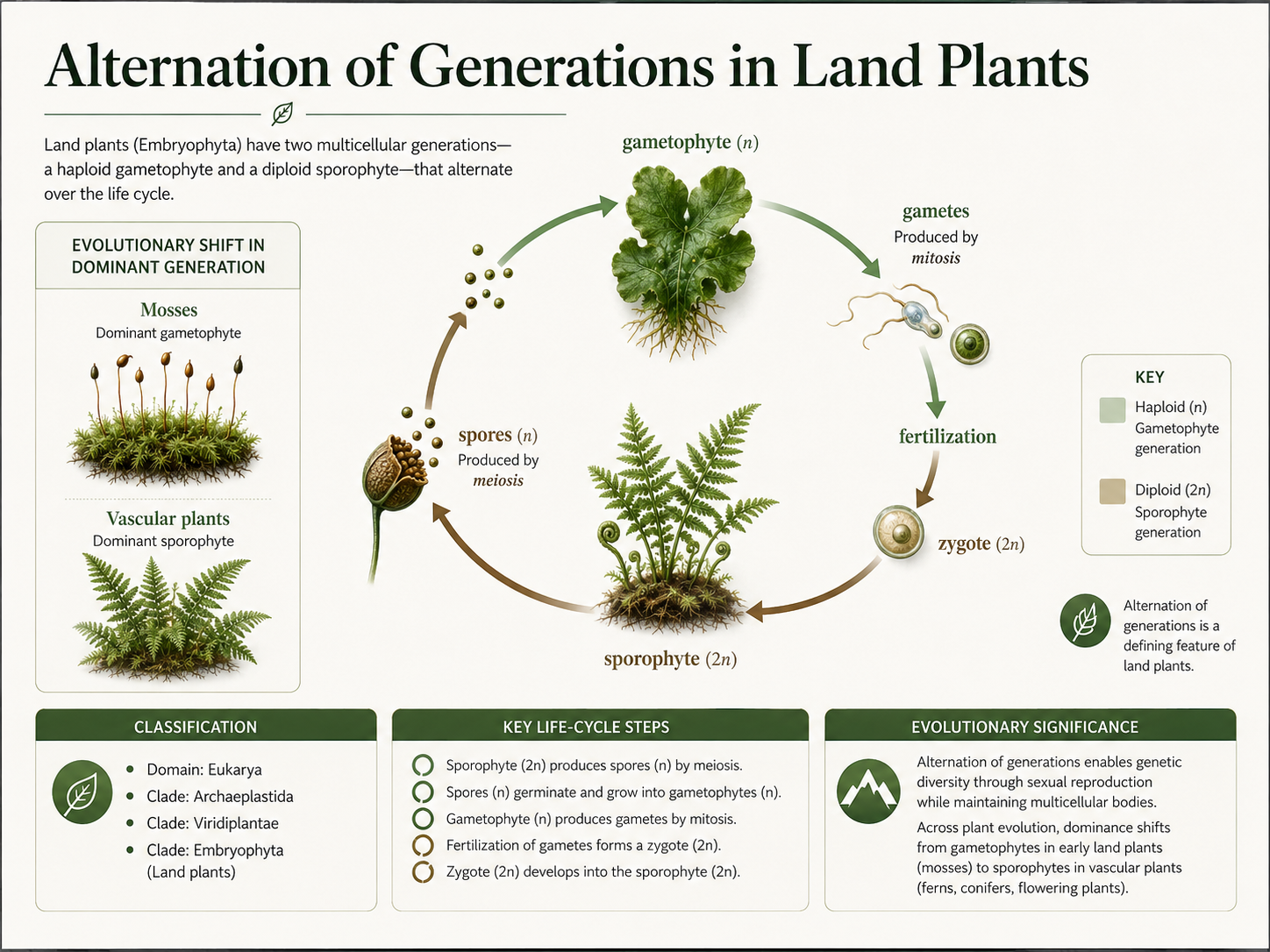
Figure 8. Alternation of Generations. Land plants alternate between a haploid gametophyte generation and a diploid sporophyte generation. The gametophyte produces gametes by mitosis, fertilization produces a diploid zygote, the zygote develops into the sporophyte, and the sporophyte produces spores by meiosis.
Nonvascular Plants: Embryophytes Without Vascular Tissue
Domain Eukarya: Archaeplastida: Viridiplantae: Embryophyta: Nonvascular Plants
The earliest-diverging living land plants are often discussed together as nonvascular plants. This group includes liverworts, mosses, and hornworts. The term is useful for teaching because these plants lack true vascular tissue: the specialized internal transport system made of xylem and phloem. However, “nonvascular plants” is a descriptive grouping, not a single clade that includes all descendants of one common ancestor while excluding none. It is better understood as a grade of early-diverging land plants that lack the vascular systems found in ferns, conifers, and flowering plants. Nonvascular plants are usually small because they do not have lignified vascular tissue for long-distance water transport and strong upright support. They absorb water and nutrients across much of their surface and often live in moist environments. Many attach to surfaces using rhizoids, which are root-like anchoring structures, but rhizoids are not true roots because they lack vascular tissue. Nonvascular plants have a gametophyte-dominant life cycle. The green, visible plant body is usually the haploid gametophyte. The diploid sporophyte remains attached to the gametophyte and depends on it nutritionally, at least partly. The dominant plant body changed during plant evolution, from gametophyte dominance in early-diverging land plants to sporophyte dominance in vascular plants.

Figure 9. Nonvascular Plants. Liverworts, mosses, and hornworts are early-diverging land plants that lack true vascular tissue. They are usually small, depend on moist environments, attach using rhizoids, and have gametophyte-dominant life cycles.
Phylum Marchantiophyta: Liverworts
Domain Eukarya: Archaeplastida: Viridiplantae: Embryophyta: Phylum Marchantiophyta
Liverworts belong to Phylum Marchantiophyta. They are among the earliest-diverging living lineages of land plants. Many liverworts grow as flattened, ribbon-like bodies called thalli, although some have small leaf-like structures. They often live in moist soils, on rocks, on tree bark, or in shaded habitats where drying stress is reduced. Liverworts lack true roots, stems, leaves, xylem, and phloem. They attach to their substrate using rhizoids. Their gametophyte is the dominant stage, and the sporophyte is usually small and dependent on the gametophyte. Like other nonvascular plants, liverworts reproduce using spores, but fertilization still requires water because sperm must swim to the egg. Liverworts matter because they show how small, simple land plants can survive without vascular tissue. Their body plan helps illustrate an early solution to terrestrial life: stay small, remain close to moisture, and protect reproductive cells in simple multicellular structures.

Figure 10. Phylum Marchantiophyta. Liverworts are small nonvascular land plants with gametophyte-dominant life cycles. Many have flattened thalli, attach using rhizoids, and live in moist environments where swimming sperm can reach eggs during reproduction.
Phylum Bryophyta: Mosses
Domain Eukarya: Archaeplastida: Viridiplantae: Embryophyta: Phylum Bryophyta
Mosses belong to Phylum Bryophyta. They are nonvascular plants that often grow in dense mats or carpets on soil, rocks, tree trunks, and other moist surfaces. The familiar green moss plant is the haploid gametophyte. It may have small leaf-like structures, known as microphylls, but these are not true leaves because they lack the same vascular organization found in vascular plants. Mosses anchor themselves using rhizoids. Because they lack true vascular tissue, they are limited in height and depend heavily on capillary movement of water across and between cells. The sporophyte grows attached to the gametophyte and usually consists of a stalk and capsule. Inside the capsule, meiosis produces spores that can disperse and grow into new gametophytes. Mosses play major ecological roles despite their small size. They help retain moisture, stabilize soil, contribute to nutrient cycles, and provide microhabitats for small organisms. Peat mosses, often placed in the genus Sphagnum, can form peatlands that store large amounts of carbon.
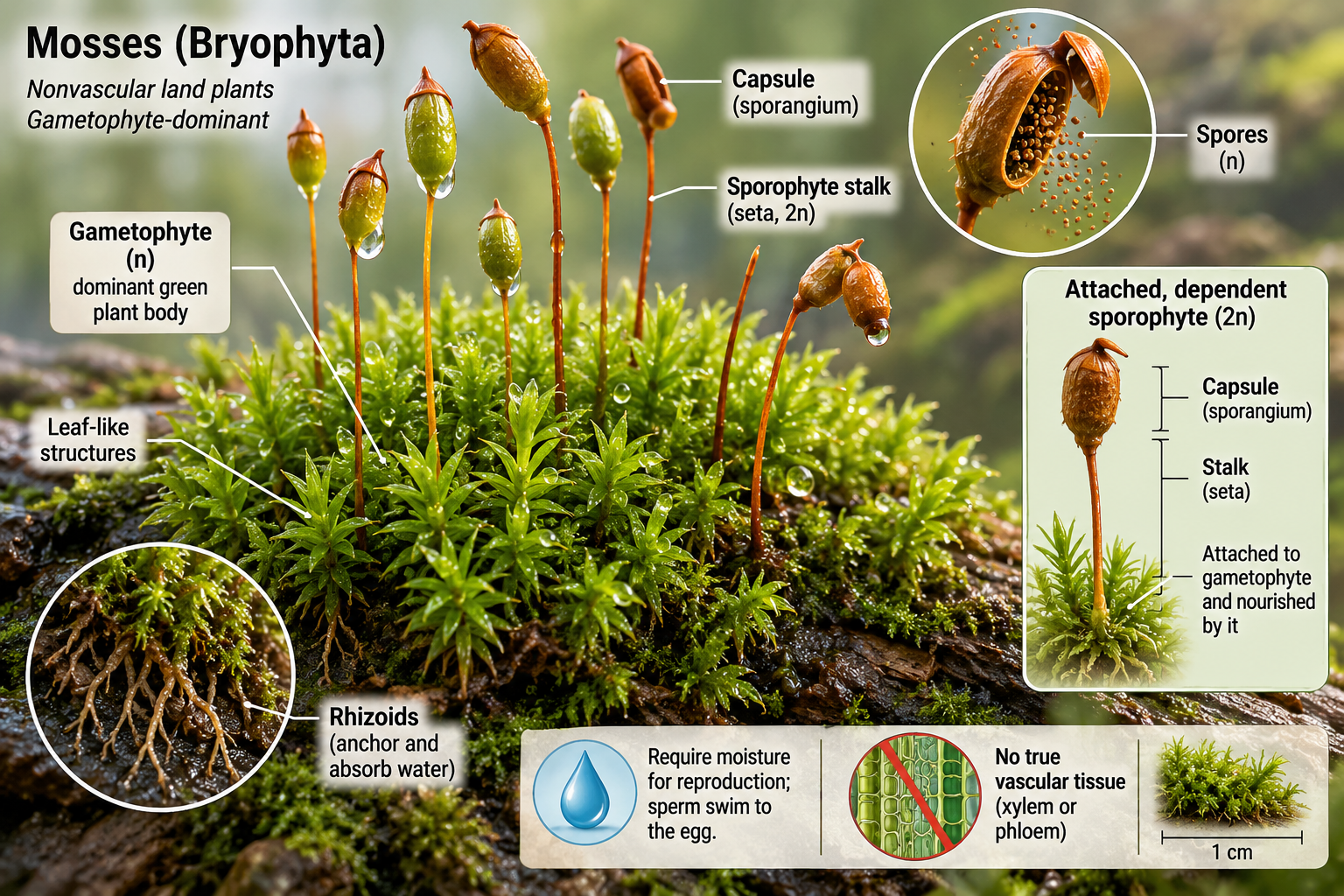
Figure 11. Phylum Bryophyta. Mosses are nonvascular land plants with a dominant gametophyte generation. They grow in mats, attach with rhizoids, and produce attached sporophytes with capsules that release spores. Their small size reflects the absence of true vascular tissue.
Phylum Anthocerotophyta: Hornworts
Domain Eukarya: Archaeplastida: Viridiplantae: Embryophyta: Phylum Anthocerotophyta
Hornworts are non-vascular plants belonging to Phylum Anthocerotophyta. Their gametophytes are usually flattened and can resemble some liverworts. Their name comes from the long, horn-like sporophytes that grow upward from the gametophyte surface. Hornwort sporophytes are unusual among nonvascular plants because they can continue growing from a basal region for an extended period. Many hornworts also have internal cavities that house symbiotic cyanobacteria. These cyanobacteria can fix nitrogen, converting atmospheric nitrogen into forms that living organisms can use. Hornworts are important because they show another version of the early land plant body plan. Like liverworts and mosses, they lack true vascular tissue and have a gametophyte-dominant life cycle. However, their distinctive sporophytes and symbioses show that early-diverging land plants are not all the same. They represent separate living lineages with their own evolutionary histories.

Figure 12. Phylum Anthocerotophyta. Hornworts are nonvascular land plants with flattened gametophytes and elongated horn-like sporophytes. Some hornworts form symbiotic relationships with nitrogen-fixing cyanobacteria.
Vascular Plants: The Origin of Internal Transport
Domain Eukarya: Archaeplastida: Viridiplantae: Embryophyta: Clade Tracheophyta
Vascular plants belong to Clade Tracheophyta. Their defining feature is vascular tissue: specialized internal tissue that transports water, minerals, sugars, and other materials through the plant body. The two major vascular tissues are xylem and phloem. Xylem transports water and dissolved minerals, usually from roots upward into stems and leaves. Xylem cells often contain lignin, a tough structural polymer that strengthens cell walls. Lignin allowed plants to grow taller because it provided support against gravity. Phloem transports sugars and other organic molecules from photosynthetic tissues to parts of the plant that need energy, such as roots, growing shoots, and developing reproductive structures. The origin of vascular tissue transformed plant evolution. Plants could grow taller, compete more effectively for light, move water from soil to aerial tissues, and support larger leaves and branching bodies. This innovation helped shift the dominant plant body from the gametophyte to the sporophyte. In vascular plants, the large visible plant is usually the diploid sporophyte.
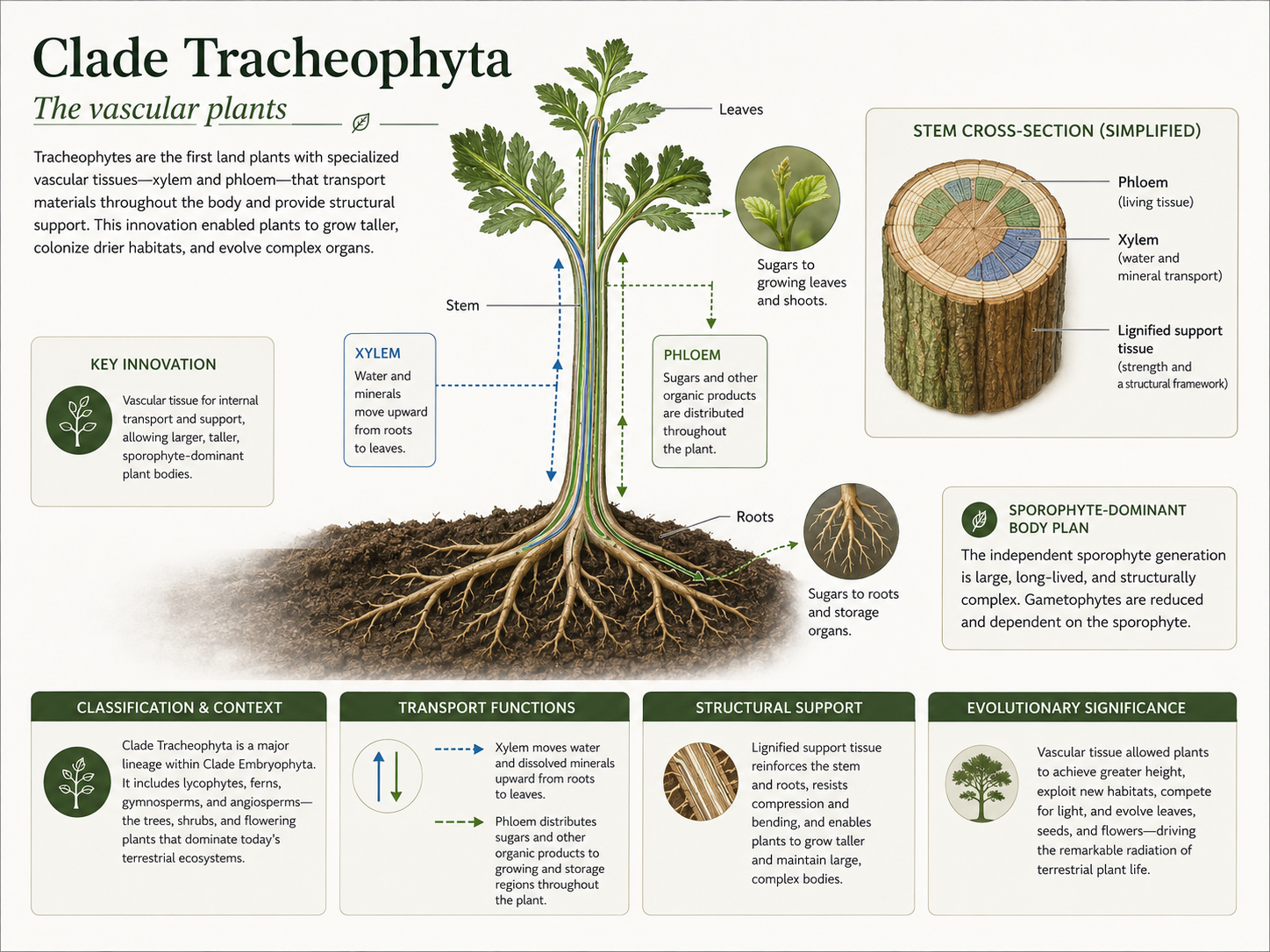
Figure 13. Origin of Vascular Tissue. Vascular plants have xylem and phloem. Xylem transports water and minerals and provides structural support through lignified cell walls. Phloem transports sugars and other organic compounds. These tissues allowed plants to grow taller and develop larger, more complex bodies.
Xylem, Phloem, Roots, and Leaves
Domain Eukarya: Archaeplastida: Viridiplantae: Embryophyta: Tracheophyta
The vascular plant body is organized around transport. Roots absorb water and minerals from soil and anchor the plant. Stems support the plant body and connect roots to leaves. Leaves increase surface area for photosynthesis. These organs work together as an integrated system. Rhizoids and true roots differ in structure and function. Rhizoids are simple anchoring structures found in nonvascular plants and some plant life stages. They can absorb water across their surface, but they lack xylem, phloem, root caps, and complex branching tissues. Their advantage is simplicity: they work well for small plants or gametophytes living close to moist surfaces. True roots are specialized vascular organs connected to the stem. They absorb water and minerals, provide stronger anchorage, explore larger soil volumes, and often store food. This helped plants become larger and less dependent on constantly moist surfaces. Early xylem was made mainly of tracheids, long narrow cells that conduct water and provide support. All vascular plants have tracheids. Some lineages later evolved vessel elements, which are wider, shorter conducting cells that move water more efficiently. Vessel elements are especially characteristic of flowering plants, although similar cells also occur in gnetophytes. Leaves also evolved in more than one way. Microphylls are small leaves with a single vascular strand and are characteristic of lycophytes. They are simple and inexpensive to build, but have limited surface area and venation. Megaphylls are larger leaves with branching veins and occur in ferns, gymnosperms, and flowering plants. Their advantage is greater light capture and photosynthetic output, but they require stronger vascular systems to supply water and remove sugars. Together, true roots, stems, xylem, phloem, and leaves allowed vascular plants to grow larger and occupy a wider range of terrestrial environments.
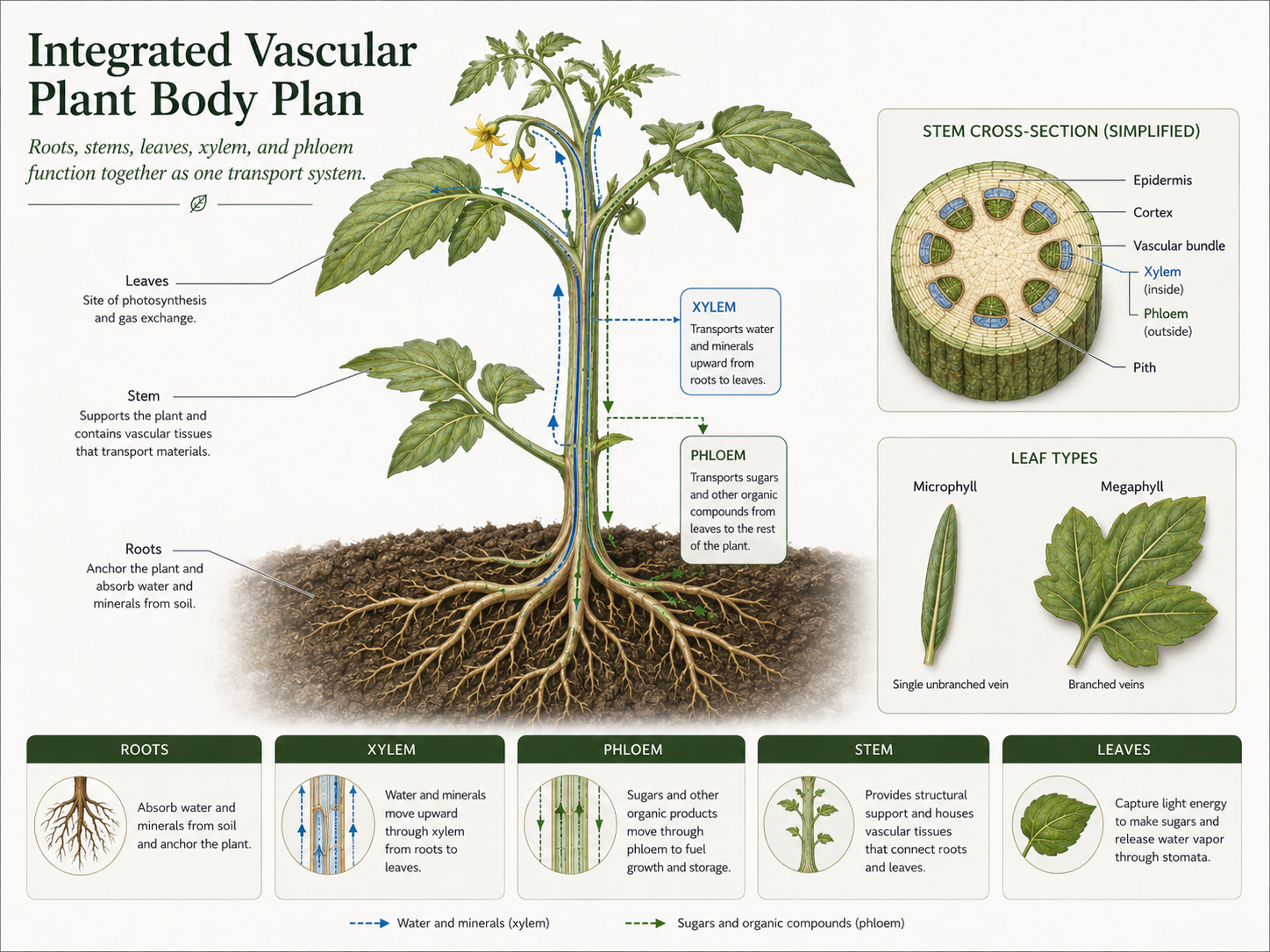
Figure 14. Vascular Plant Organs. Roots absorb water and minerals, stems support and connect the plant body, and leaves capture light for photosynthesis. Xylem and phloem link these organs into a single transport system.
Lycophytes: Club Mosses and Their Relatives
Domain Eukarya: Archaeplastida: Viridiplantae: Embryophyta: Tracheophyta: Phylum Lycopodiophyta
Lycophytes belong to Phylum Lycopodiophyta and represent one of the earliest-diverging living lineages of vascular plants. Living lycophytes include club mosses, spike mosses, and quillworts. Despite the common name “club moss,” lycophytes are not true mosses. They are vascular plants with xylem and phloem. Lycophytes have microphylls, small leaves with a single vein. This distinguishes them from ferns and seed plants, which generally have megaphylls with branching veins. Many lycophytes produce spores in structures associated with specialized leaves. In some species, these spore-producing leaves are grouped into cone-like structures called strobili. Lycophytes are small today, but their ancient relatives were once major components of Carboniferous forests. Some extinct lycophytes grew as large trees and contributed to the plant material that eventually formed coal deposits. Living lycophytes are modest in size, but their lineage preserves a major early branch in vascular plant evolution.

Figure 15. Phylum Lycopodiophyta. Lycophytes are vascular plants with microphylls, xylem, and phloem. Club mosses are not true mosses; they belong to an early-diverging vascular plant lineage that was once represented by large tree-like forms in ancient forests.
Ferns and Their Relatives: Seedless Vascular Plants with Megaphylls
Domain Eukarya: Archaeplastida: Viridiplantae: Embryophyta: Tracheophyta: Clade Monilophyta
True ferns and their relatives are seedless vascular plants often grouped in Clade Monilophyta. This group includes true ferns, horsetails, and whisk ferns. They have vascular tissue and dominant sporophytes, but they do not produce seeds. Like mosses and liverworts, they reproduce using spores. Unlike mosses and liverworts, the familiar fern plant is the diploid sporophyte. Seedless vascular plants still depend on water for fertilization. Their sperm are flagellated and must swim through a thin film of water to reach the egg. This is why many ferns and their relatives are common in moist, shaded environments, even though some species are adapted to drier habitats.
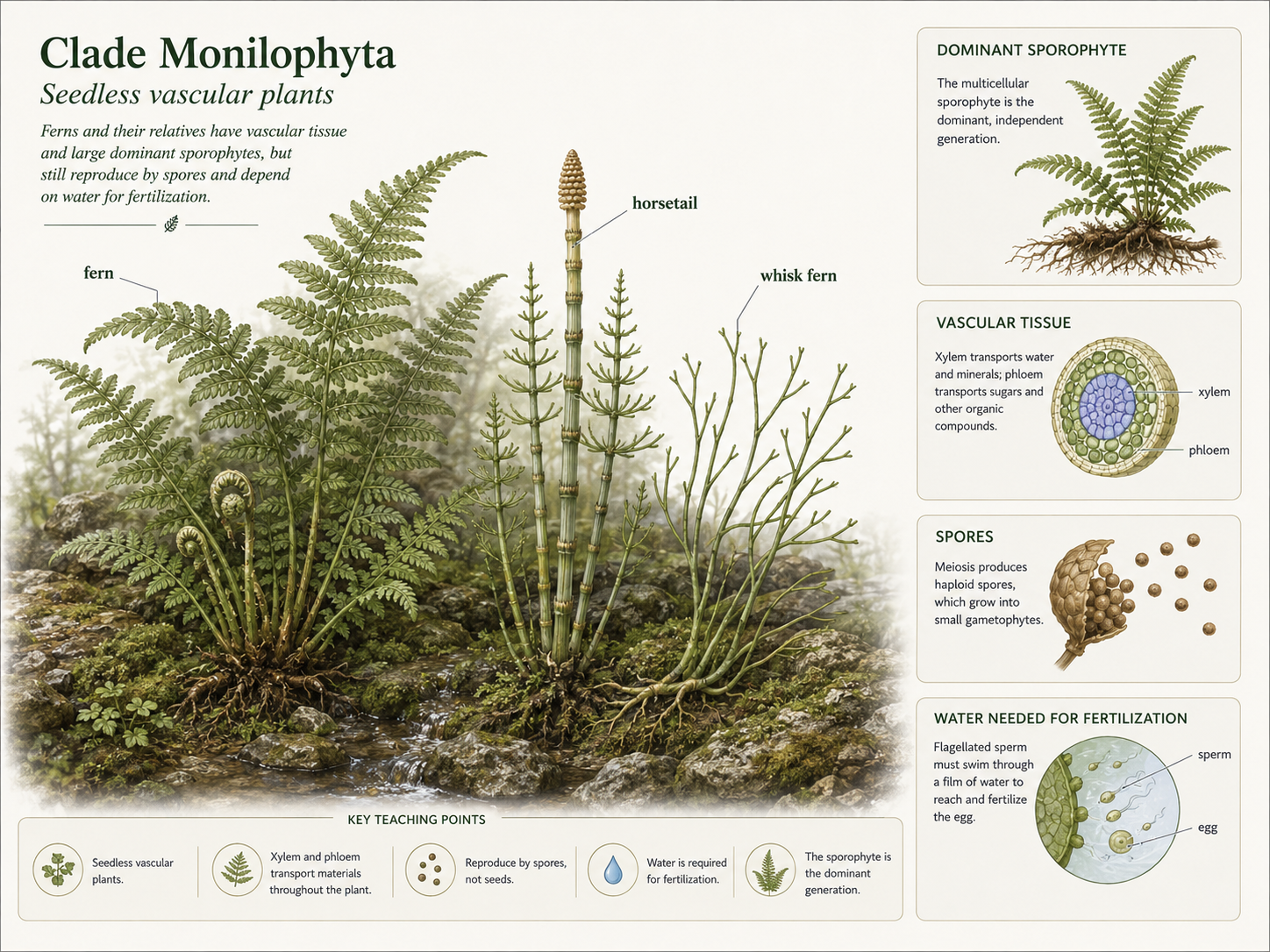
Figure 16. Seedless Vascular Plants. Ferns and their relatives have xylem, phloem, and dominant sporophytes, but they reproduce by spores rather than seeds. Their sperm require water for fertilization, linking them to moist environments.
Whisk Ferns
Domain Eukarya: Archaeplastida: Viridiplantae: Embryophyta: Tracheophyta: Monilophyta: Class Psilotopsida
Whisk ferns are unusual seedless vascular plants in Class Psilotopsida. They have simple, branching green stems and lack true roots and true leaves. Instead of leaves, they have small scale-like outgrowths called enations. Enations may look leaf-like, but they lack vascular tissue and are not considered true leaves. Whisk ferns anchor themselves with rhizome-like structures and rhizoids, often in association with mycorrhizal fungi. These fungi help the plant obtain water and nutrients. Whisk ferns are not “primitive ancestors” of other vascular plants. Their simple body plan likely reflects both ancient traits and evolutionary reduction. They are useful because they show that vascular plants can vary widely in root and leaf structure.
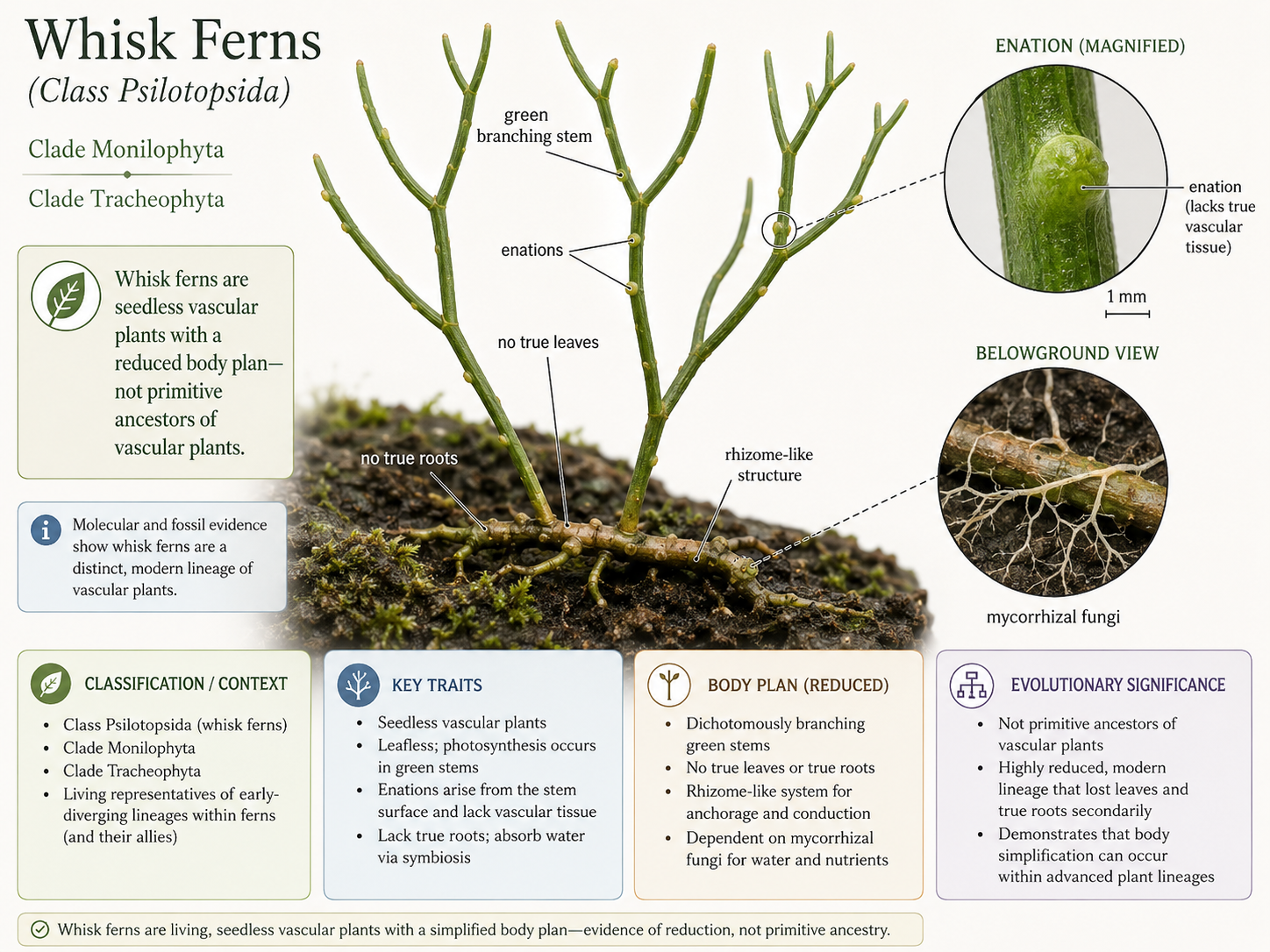
Figure 17. Class Psilotopsida. Whisk ferns are seedless vascular plants with green branching stems, no true roots, and no true leaves. Their small enations resemble leaves but lack vascular tissue.
Horsetails
Domain Eukarya: Archaeplastida: Viridiplantae: Embryophyta: Tracheophyta: Monilophyta: Class Equisetopsida
Horsetails belong to Class Equisetopsida. Living horsetails are placed in the genus Equisetum. They have jointed, ribbed stems with nodes, and their leaves are highly reduced. The leaves often form small whorls around the nodes, making the stem appear segmented. Horsetails reproduce by spores produced in cone-like structures called strobili at the tips of some stems. Their stems contain silica, which gives them a rough texture. Historically, some horsetails were called scouring rushes because people used them to scrub pots and surfaces. Horsetails are the surviving representatives of a once much more diverse lineage. Ancient horsetail relatives included larger forms that grew in wet Carboniferous landscapes. Modern horsetails are much smaller, but they preserve an important branch of seedless vascular plant diversity.

Figure 18. Class Equisetopsida. Horsetails are seedless vascular plants with jointed stems, reduced leaves, and spore-producing strobili. Their silica-rich stems and segmented appearance make them easy to recognize.
True Ferns
Domain Eukarya: Archaeplastida: Viridiplantae: Embryophyta: Tracheophyta: Monilophyta: Class Polypodiopsida
True ferns belong to Class Polypodiopsida. They are among the most familiar seedless vascular plants. The visible fern plant is the diploid sporophyte, usually with large leaves called fronds. Fern fronds are megaphylls, meaning they have branching vascular tissue. This gives ferns a larger photosynthetic surface area than plants with tiny single-veined microphylls. Many ferns produce spores in clusters called sori, usually on the underside of fronds. When spores germinate, they grow into small, independent gametophytes. These gametophytes produce sperm and eggs. Fertilization requires water, and the resulting zygote grows into a new sporophyte. Ferns show a major evolutionary step beyond nonvascular plants. They have true vascular tissue, true roots, and large leaves, but they still reproduce by spores and require water for fertilization. This combination makes them excellent examples of seedless vascular plants.
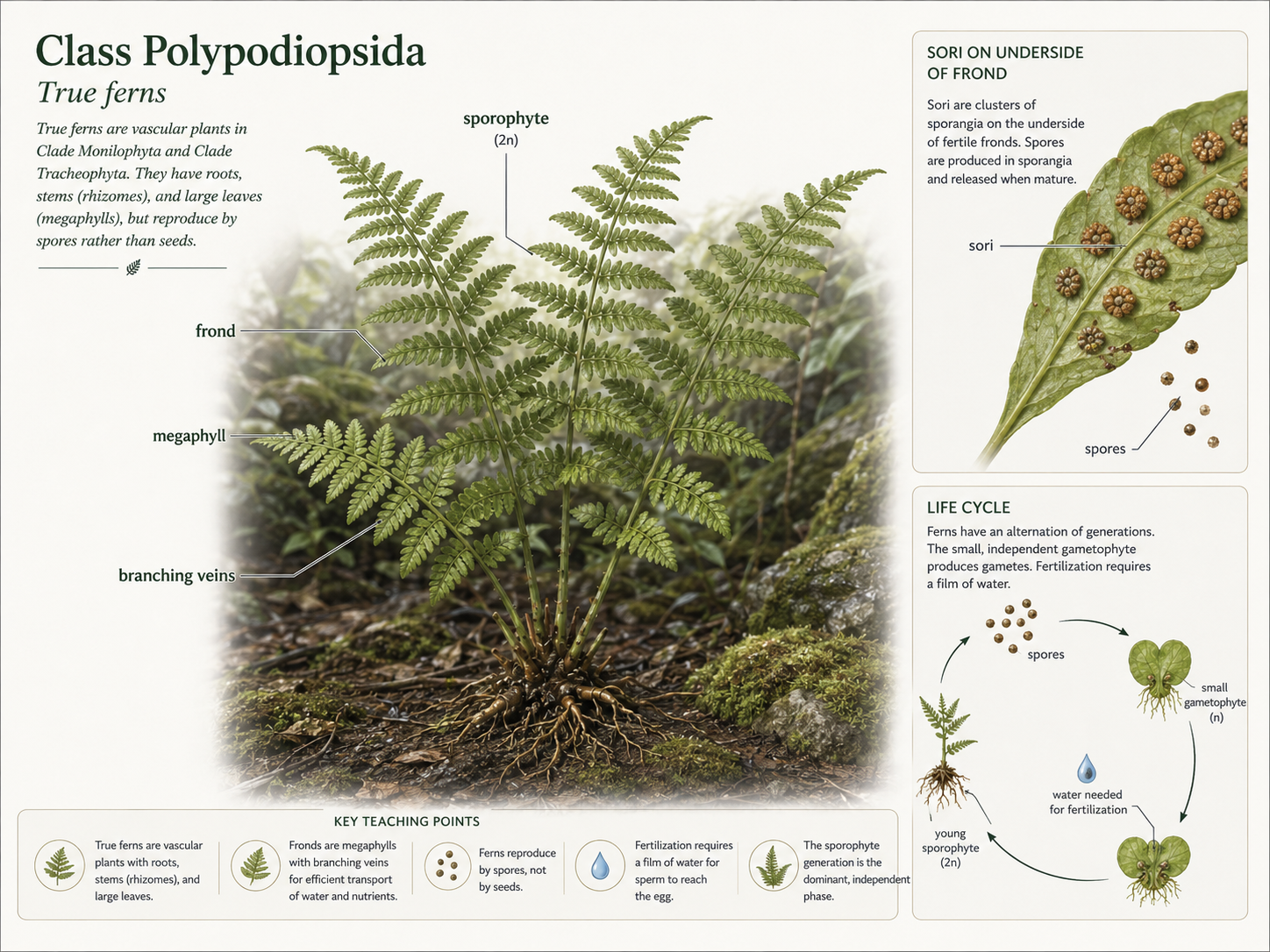
Figure 19. Class Polypodiopsida. True ferns are seedless vascular plants with roots, xylem, phloem, and large fronds called megaphylls. Spores are often produced in sori on the underside of fronds, and fertilization still requires water.
Seed Plants: The Origin of Seeds and Pollen
Domain Eukarya: Archaeplastida: Viridiplantae: Embryophyta: Tracheophyta: Clade Spermatophyta
Seed plants belong to Clade Spermatophyta. They include gymnosperms and angiosperms and are the dominant plants in many terrestrial ecosystems today. The visible seed plant is the diploid sporophyte, which produces pollen and seeds as part of its reproductive cycle. Pollen is the male gametophyte. It carries sperm cells and protects them from drying, allowing fertilization to occur without swimming sperm moving through external water. Seeds contain a plant embryo, a protective seed coat, and stored nutrients or nutritive tissue. Unlike spores, seeds protect and nourish the young plant and can remain dormant until conditions improve. Seed plants show a major evolutionary step beyond earlier seedless vascular plants. They have true vascular tissue, true roots, and reproductive structures that reduce dependence on standing water. They did not evolve from modern ferns, but likely arose from extinct vascular plant relatives with wood and heterospory. Heterospory means producing two types of spores: microspores, which lead to male gametophytes, and megaspores, which lead to female gametophytes. This combination of pollen, seeds, vascular tissue, and heterospory helped seed plants spread widely across land.

Figure 20. Origin of Seed Plants. Seed plants produce pollen and seeds. Pollen protects and transports the male gametophyte, while seeds protect, nourish, and disperse the embryo. These innovations reduced dependence on water for fertilization and helped seed plants spread across terrestrial environments.
Gymnosperms: Seed Plants with Exposed Ovules
Domain Eukarya: Archaeplastida: Viridiplantae: Embryophyta: Tracheophyta: Spermatophyta: Gymnosperm Lineages
Gymnosperms are seed plants that produce ovules and seeds that are not enclosed inside fruits. They belong within Clade Spermatophyta and include cycads, ginkgo, conifers, and gnetophytes. The visible gymnosperm plant is the diploid sporophyte, usually a woody tree or shrub. The word gymnosperm means “naked seed,” but gymnosperm seeds are not literally unprotected. They are often produced on cone scales or other exposed reproductive structures rather than enclosed inside an ovary. Many gymnosperms rely on wind to move pollen, although some cycads and gnetophytes involve insects in pollination. Gymnosperms show an important stage in seed plant evolution. They have vascular tissue, wood, pollen, and seeds, but they do not produce flowers or fruits. Gymnosperms were especially common and ecologically important in many Mesozoic landscapes before flowering plants became dominant in many environments. They remain important today. Conifers form vast forests, cycads preserve ancient-looking tropical body plans, ginkgo is the single living species of a once more diverse lineage, and gnetophytes show unusual combinations of traits.

Figure 21. Gymnosperms. Gymnosperms are seed plants with ovules and seeds not enclosed inside fruits. Living gymnosperms include cycads, ginkgo, conifers, and gnetophytes. They reproduce using pollen and seeds and include many woody plants adapted to terrestrial environments.
Class Cycadopsida: Cycads
Domain Eukarya: Archaeplastida: Viridiplantae: Embryophyta: Tracheophyta: Spermatophyta: Gymnosperms: Class Cycadopsida
Cycads are gymnosperms in Class Cycadopsida. They are seed plants with thick trunks, large stiff compound leaves, and cone-like reproductive structures. The visible cycad plant is the diploid sporophyte. Many cycads superficially resemble palms, but they are not palms. Palms are flowering plants in the angiosperm lineage, while cycads are gymnosperms. Cycads produce seeds, but those seeds are not enclosed inside fruits. Many species have separate male and female plants, with pollen produced by male plants and ovules produced by female plants. Cycads show an important example of gymnosperm diversity. They have vascular tissue, wood, pollen, and seeds, but they do not produce flowers or fruits. Although cycads are often described as ancient-looking, living cycads are not unchanged fossils. They are modern organisms with their own evolutionary history, and some living cycad lineages diversified relatively recently. Cycads are most common in tropical and subtropical regions, and many species are threatened by habitat loss, slow growth, and illegal collection. This combination of ancient seed plant features and modern conservation concern makes cycads important both evolutionarily and biologically.

Figure 22. Class Cycadopsida. Cycads are gymnosperms with thick trunks, large compound leaves, and cone-like reproductive structures. They may resemble palms, but palms are flowering plants, while cycads belong to a separate seed plant lineage.
Class Ginkgoopsida: Ginkgo
Domain Eukarya: Archaeplastida: Viridiplantae: Embryophyta: Tracheophyta: Spermatophyta: Gymnosperms: Class Ginkgoopsida
Ginkgo belongs to Class Ginkgoopsida. Today this gymnosperm lineage is represented by a single living species, Ginkgo biloba. Ginkgo trees are seed plants with distinctive fan-shaped leaves, woody stems, and seeds with fleshy outer coverings. The visible ginkgo tree is the diploid sporophyte. Ginkgo is deciduous, meaning it loses its leaves seasonally. Like other gymnosperms, it produces seeds, but those seeds are not enclosed inside fruits. Ginkgo shows an important example of a once more diverse seed plant lineage that is now represented by only one living species. It is often called a living fossil because fossils with ginkgo-like leaves are known from deep time, and the living species resembles ancient relatives in some visible traits. However, this phrase should be used carefully. Living ginkgo trees are not frozen in time. They are modern organisms that continued evolving while retaining some recognizable ancient features. Ginkgo survived in cultivation for centuries and is widely planted in cities today because it tolerates pollution, compacted soils, and other urban stresses. Its wild status is complicated because some populations may reflect ancient cultivation and later naturalization.

Figure 23. Class Ginkgoopsida. Ginkgo biloba is the only living species of a once more diverse gymnosperm lineage. It has fan-shaped leaves, exposed seeds, and a long fossil history, but living ginkgo trees are modern plants rather than unchanged relics.
Class Pinopsida: Conifers
Domain Eukarya: Archaeplastida: Viridiplantae: Embryophyta: Tracheophyta: Spermatophyta: Gymnosperms: Class Pinopsida
Conifers are gymnosperms in Class Pinopsida. This group includes pines, spruces, firs, cedars, cypresses, junipers, redwoods, and their relatives. Most conifers are seed plants with woody stems and needle-like or scale-like leaves. The visible conifer plant is the diploid sporophyte, usually a tree or shrub. Many conifers are evergreen, meaning they keep their leaves for more than one growing season. However, some conifers, such as bald cypress and larch, are deciduous and lose their leaves seasonally. Conifers usually produce seeds in cones. Male cones produce pollen, and female cones bear ovules that develop into seeds after fertilization. Like other gymnosperms, conifers produce seeds, but those seeds are not enclosed inside fruits. Many conifers rely on wind pollination and produce large amounts of pollen that can travel through the air. Conifers show one of the most ecologically important branches of gymnosperm diversity. They have vascular tissue, wood, pollen, and seeds, but they do not produce flowers or fruits. Conifers dominate vast boreal forests, form mountain forests, and include some of the tallest and oldest living trees. Coast redwoods, Sequoia sempervirens, are the tallest living trees, and Great Basin bristlecone pines, Pinus longaeva, include some of the oldest known individual trees.

Figure 24. Class Pinopsida. Conifers are woody gymnosperms that usually have needle-like or scale-like leaves and seeds borne in cones. They include pines, spruces, firs, cypresses, junipers, redwoods, and some of the tallest and oldest living trees.
Class Gnetopsida: Gnetophytes
Domain Eukarya: Archaeplastida: Viridiplantae: Embryophyta: Tracheophyta: Spermatophyta: Gymnosperms: Class Gnetopsida
Gnetophytes are gymnosperms in Class Gnetopsida. This group includes three living genera: Gnetum, Ephedra, and Welwitschia. The visible gnetophyte plant is the diploid sporophyte, but these plants look dramatically different from one another. Gnetum includes tropical trees, shrubs, and vines with broad leaves. Ephedra includes desert shrubs with jointed green stems. Welwitschia mirabilis, found in the Namib Desert region of southwestern Africa, produces only two main leaves that continue growing throughout the life of the plant. Like other gymnosperms, gnetophytes produce seeds, but those seeds are not enclosed inside fruits. Gnetophytes are especially interesting because they share some traits with flowering plants, including vessel-like water-conducting cells and, in some cases, insect-associated pollination. Earlier botanists used these similarities to suggest that gnetophytes might be close relatives or even ancestors of flowering plants. Modern genetic evidence does not support the idea that flowering plants evolved from living gnetophytes. Instead, these similarities are usually interpreted as convergent evolution, meaning similar traits evolved independently in separate lineages. Gnetophytes show that gymnosperms are not a single uniform body type, but a diverse group of seed plants with very different forms and ecological strategies.

Figure 25. Class Gnetopsida. Gnetophytes are unusual gymnosperms that include Gnetum, Ephedra, and Welwitschia. Some gnetophytes share superficial similarities with flowering plants, such as vessel-like conducting cells and insect-associated pollination, but these traits do not mean that angiosperms evolved from living gnetophytes.
Angiosperms: Flowering Plants
Domain Eukarya: Archaeplastida: Viridiplantae: Embryophyta: Tracheophyta: Spermatophyta: Clade Angiospermae
Angiosperms are flowering plants in Clade Angiospermae. They are seed plants that produce flowers and fruits and are the most diverse living plant group. Angiosperms include grasses, lilies, orchids, oaks, roses, beans, cacti, sunflowers, maples, palms, and most crop plants. The visible angiosperm plant is the diploid sporophyte. Like other seed plants, angiosperms produce pollen and seeds, but they differ from gymnosperms because their seeds develop inside ovaries that mature into fruits. A flower is a reproductive shoot with specialized structures that produce and protect gametophytes, support fertilization, and often attract or interact with pollinators. Flowers vary enormously. Some are large, colorful, and animal-pollinated, while others are small, reduced, and wind-pollinated, as in many grasses. A fruit is a mature ovary or associated floral tissue that encloses seeds. Fruits protect seeds and often help with seed dispersal. Not all fruits are sweet or fleshy. Pea pods, acorns, grains, maple samaras, and peanut shells are fruits in the botanical sense because they develop from flowers and enclose seeds. Angiosperms show a major evolutionary step in seed plant history. Flowers and fruits improved pollination efficiency, seed protection, and seed dispersal, helping flowering plants diversify across many terrestrial ecosystems. Many angiosperms also form close ecological relationships with animals, fungi, and microbes, especially through animal pollination and animal-mediated seed dispersal.

Figure 26. Angiosperms. Angiosperms are flowering plants that produce seeds enclosed within fruits. Flowers support pollination and fertilization, while fruits protect seeds and often aid dispersal. This combination helped flowering plants become the most diverse living plant lineage.
The Origin and Diversification of Flowering Plants
Domain Eukarya: Archaeplastida: Viridiplantae: Embryophyta: Tracheophyta: Spermatophyta: Angiospermae
Angiosperms are flowering plants in Clade Angiospermae. They are seed plants within Clade Spermatophyta, but they did not evolve from any living gymnosperm group. Angiosperms share ancestry with other seed plants, and their exact extinct relatives remain an active area of research. The origin of flowering plants was once called an “abominable mystery” because angiosperms appear relatively suddenly and diversely in the fossil record compared with some earlier plant groups. Older hypotheses suggested that angiosperms might be closely related to gnetophytes because both groups have vessel-like conducting cells and some insect-associated pollination. Modern molecular evidence does not support that simple interpretation. Instead, these similarities are usually interpreted as convergent evolution, meaning similar traits evolved independently in separate lineages. Efficient water transport and animal-associated pollination evolved more than once in seed plant history. Modern flowering plant classification is strongly influenced by molecular phylogenetics. The Angiosperm Phylogeny Group system recognizes major lineages such as early-diverging angiosperms, magnoliids, monocots, and eudicots. This is more accurate than simply dividing all flowering plants into “monocots” and “dicots.” Angiosperms show that evolutionary history is not always obvious from visible traits alone. Molecular evidence helps reveal relationships that older classifications based only on morphology could miss.
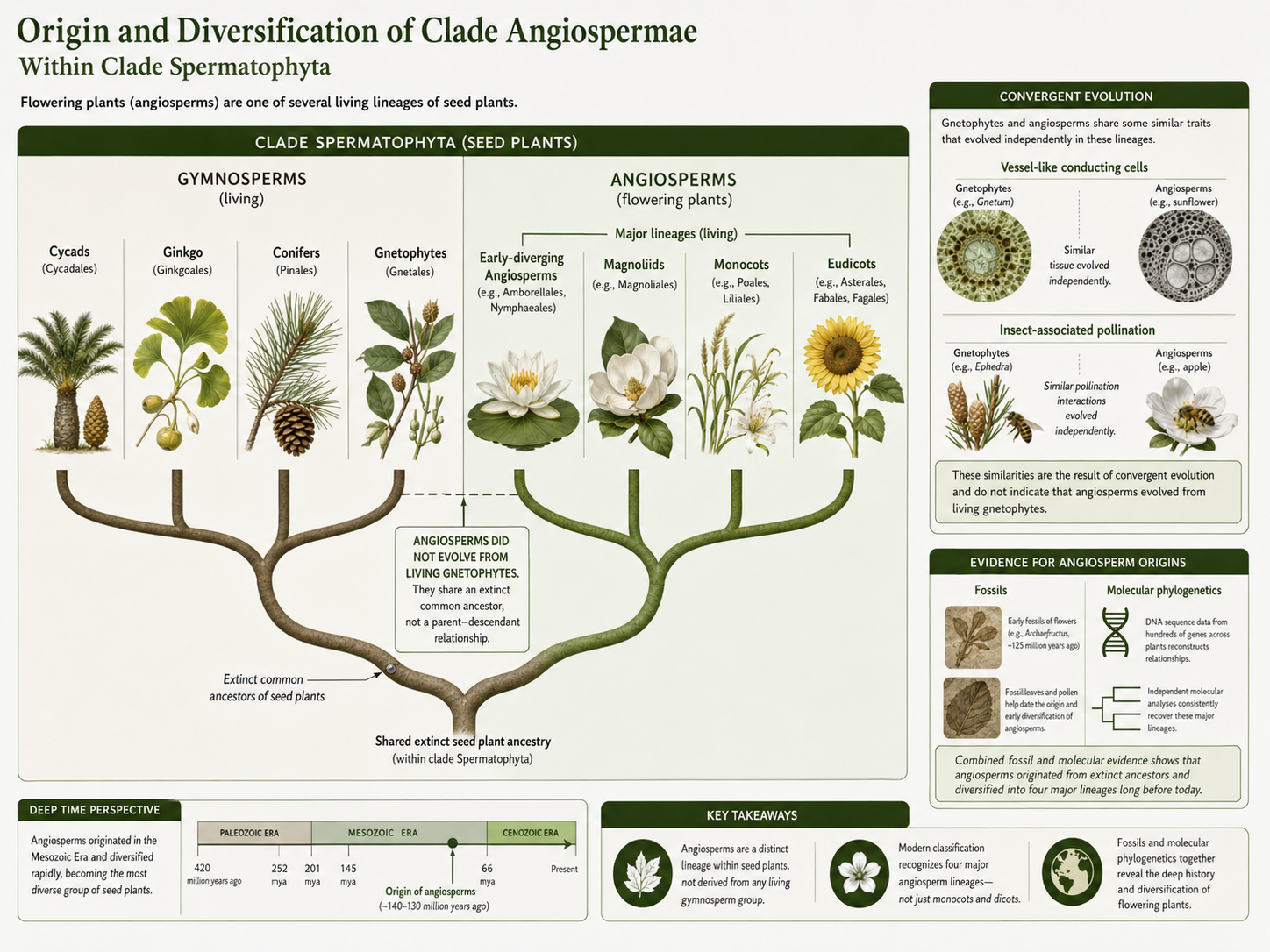
Figure 27. Origin of Flowering Plants. Angiosperms are seed plants with flowers and fruits. They did not evolve from living gnetophytes, even though some gnetophytes and angiosperms share similar traits. Modern angiosperm classification is based heavily on molecular evidence and recognizes several major flowering plant lineages.
Eudicots and the Problem with “Dicots”
Domain Eukarya: Archaeplastida: Viridiplantae: Embryophyta: Tracheophyta: Spermatophyta: Angiospermae: Clade Eudicots
Eudicots are flowering plants within Clade Angiospermae. They are one of the major modern angiosperm lineages and include many familiar plants, such as roses, oaks, beans, sunflowers, maples, and dandelions. Traditional biology courses often divided flowering plants into monocots and dicots. This comparison is useful for learning common plant traits, but it is not fully accurate as a modern evolutionary classification. “Dicot” means a flowering plant with two cotyledons, or seed leaves. Many flowering plants have two cotyledons, but not all of them belong to a single exclusive clade. The more precise modern term is eudicots, meaning “true dicots.” Eudicots typically have two cotyledons, branching leaf venation, flower parts often in fours or fives, and pollen with three openings or grooves. These traits are useful patterns, but they are not perfect rules for identifying every species. Eudicots show a major branch of flowering plant diversity. They include herbs, shrubs, trees, vines, aquatic plants, desert plants, carnivorous plants, and many crop species. Their success reflects the broader angiosperm pattern: flexible growth forms, diverse flowers, protected seeds, and many ecological partnerships.

Figure 28. Eudicots. Eudicots are a major clade of flowering plants traditionally included within “dicots.” They often have two cotyledons, branching leaf veins, flower parts in fours or fives, and tricolpate pollen. The older term “dicot” is useful descriptively but can be misleading phylogenetically.
Monocots
Domain Eukarya: Archaeplastida: Viridiplantae: Embryophyta: Tracheophyta: Spermatophyta: Angiospermae: Clade Monocots
Monocots are flowering plants within Clade Angiospermae. They are one of the major modern angiosperm lineages and include grasses, lilies, orchids, irises, palms, bananas, sedges, rushes, and many aquatic plants. The name monocot refers to the presence of one cotyledon, or seed leaf, in the embryo. Monocots usually have parallel leaf venation, flower parts in multiples of three, scattered vascular bundles in the stem, and fibrous root systems. These traits are useful patterns for introductory identification, but they are not perfect rules for every species. Monocots show a major branch of flowering plant diversity. They include herbs, aquatic plants, climbers, and large tree-like forms such as palms and bananas, although these do not produce true wood in the same way many eudicot trees do. Monocots are especially important ecologically and economically. Grasses, which belong to Family Poaceae within the monocots, form grasslands and include major human food crops such as rice, wheat, corn, barley, oats, and sugarcane. Orchids are one of the largest flowering plant families, and palms are important in tropical ecosystems and human economies. This combination of distinctive body plans, diverse habitats, and major food crops makes monocots one of the most important flowering plant groups.

Figure 29. Monocots. Monocots are flowering plants with one cotyledon. They often have parallel leaf venation, flower parts in threes, fibrous roots, and scattered vascular bundles. The group includes grasses, lilies, orchids, palms, and many major crop plants.
Plant Evolution as a Sequence of Innovations
The history of plants can be understood as a series of evolutionary innovations layered over time. Primary endosymbiosis produced plastids and opened the door to photosynthetic eukaryotes. Green plants evolved chlorophyll b and cellulose-rich cell walls. Streptophyte algae gave rise to the lineage that eventually produced land plants. Land plants evolved protected embryos, cuticles, stomata, protected reproductive structures, and resistant spores. Vascular plants evolved xylem, phloem, lignified support, roots, and leaves. Seed plants evolved pollen and seeds, reducing dependence on water for reproduction. Angiosperms evolved flowers and fruits, creating new possibilities for pollination, seed protection, and dispersal. This sequence should not be read as a ladder of progress from “simple” to “advanced.” Living liverworts, mosses, ferns, conifers, and flowering plants are all modern organisms. Each has continued evolving. The point is not that one living group became another living group. The point is that major traits appeared at different points in plant evolutionary history, and those traits opened new ecological possibilities.
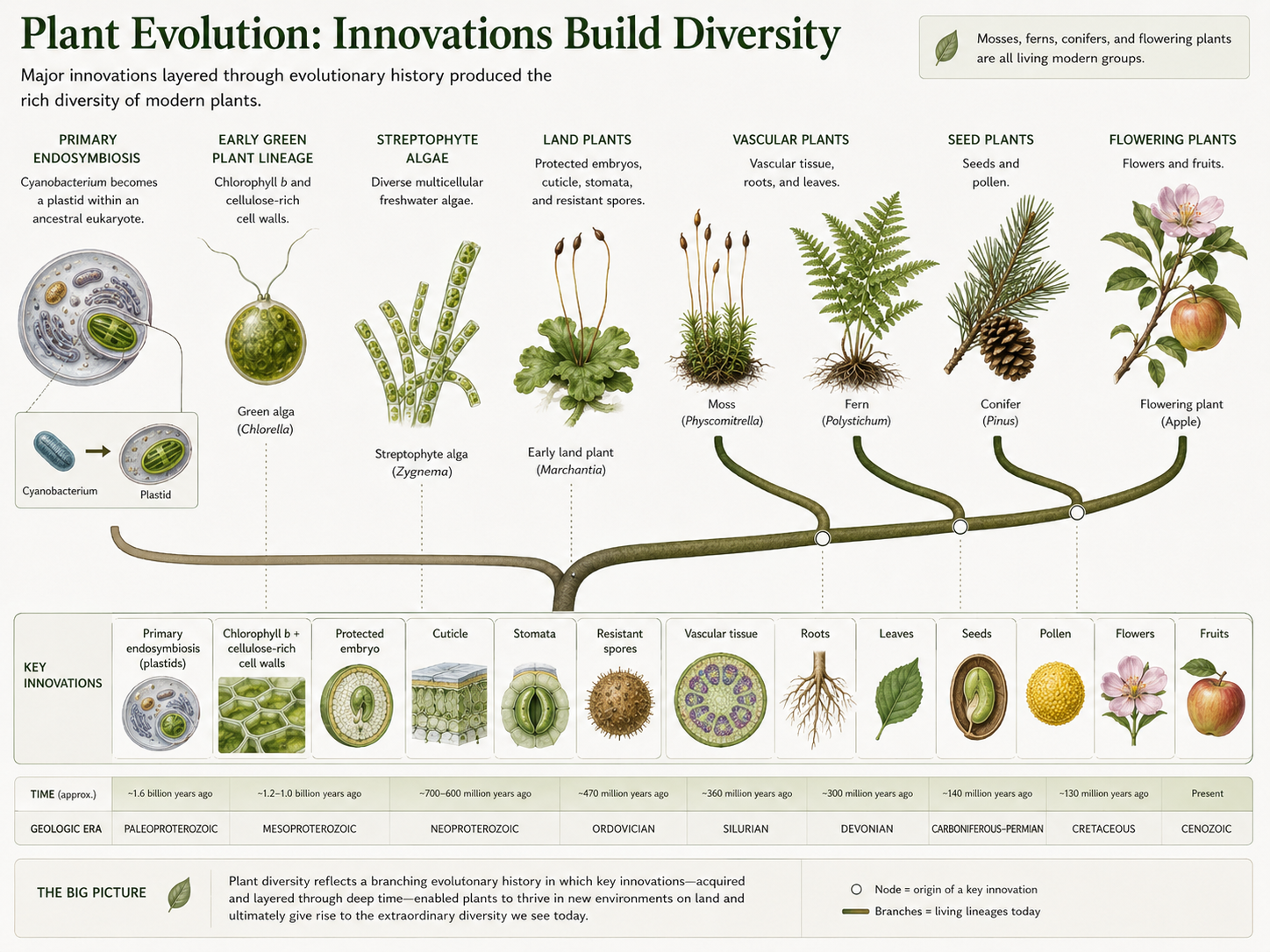
Figure 30. Major Innovations in Plant Evolution. Plant history includes the origin of primary plastids, chlorophyll b, multicellularity, protected embryos, cuticles, stomata, vascular tissue, roots, leaves, seeds, pollen, flowers, and fruits. These innovations allowed plants to diversify from aquatic photosynthetic eukaryotes into the dominant primary producers of many terrestrial ecosystems.