
Chapter: Fungi
Characteristics of Fungi
Kingdom Fungi
Fungi are eukaryotic heterotrophs that obtain nutrients by external digestion and absorption. Like animals, fungi are chemoorganoheterotrophs, meaning they obtain energy and carbon from organic molecules, but animals and fungi process food in different ways. Animals usually ingest food and digest it inside the body, while fungi release digestive enzymes into their surroundings and absorb the smaller molecules that result. This feeding strategy allows fungi to grow through soil, wood, dead organisms, living hosts, and plant roots while absorbing nutrients directly from the environment. Most fungi are multicellular, but some are unicellular. Yeasts are unicellular fungi, while molds, mushrooms, shelf fungi, puffballs, and many plant-associated fungi are multicellular. The body of a multicellular fungus is usually made of thin branching filaments called hyphae. Together, these hyphae form a larger network called a mycelium, which is the main feeding body of the fungus. A mushroom is only the reproductive structure, or fruiting body, produced by some fungi. All fungi share two major traits: they absorb nutrients after external digestion, and their cell walls contain chitin, a tough polysaccharide that provides support and protection. Chitin is also found in arthropod exoskeletons.
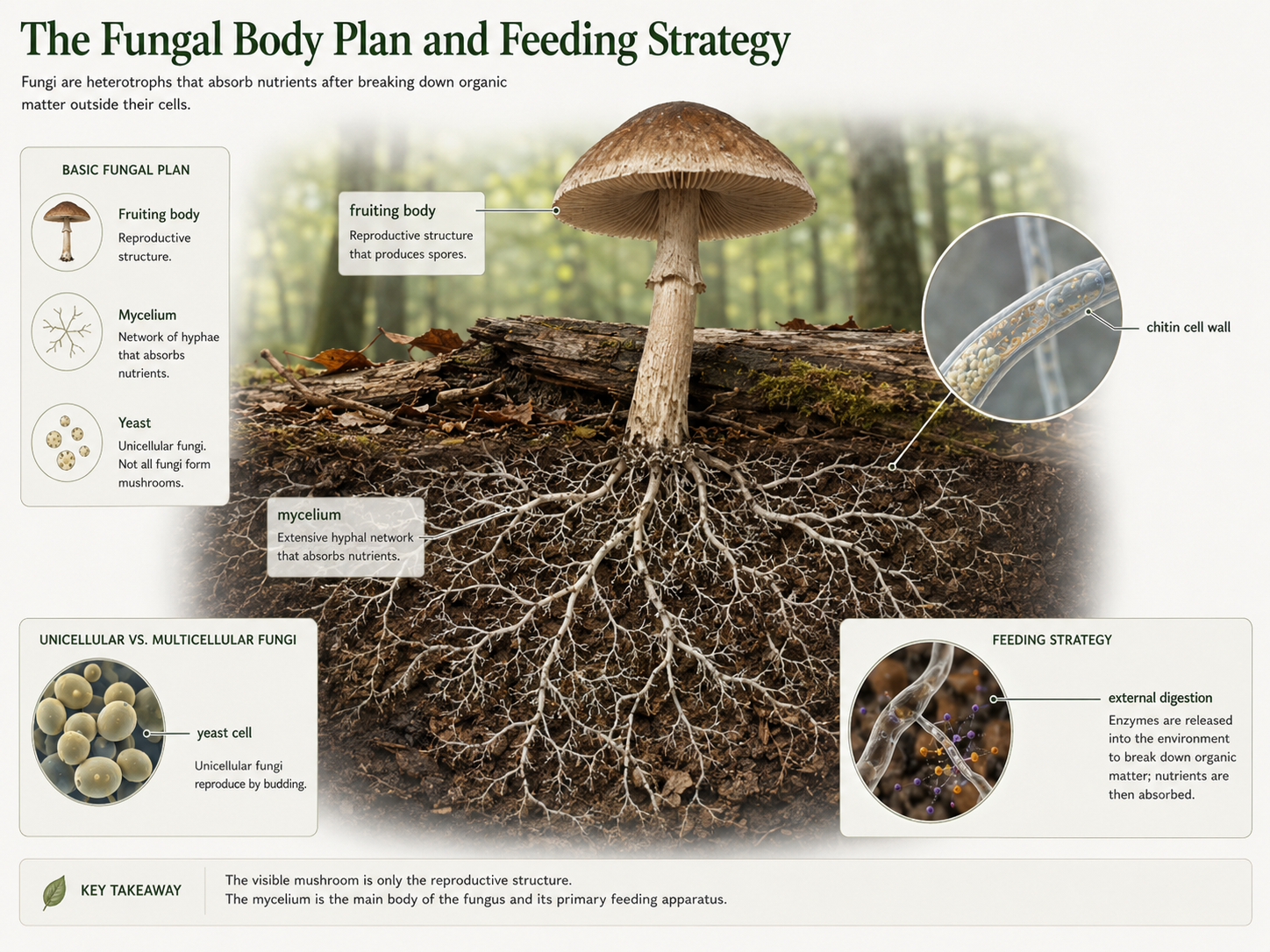
Figure 1. Characteristics of Fungi. Fungi are absorptive heterotrophs with chitin-containing cell walls. Many fungi grow as branching hyphae that form a mycelium, while yeasts are unicellular. Fungi digest food externally by releasing enzymes into the environment and absorbing the resulting nutrients.
Opisthokonta: The Evolutionary Context of Fungi
Domain Eukarya: Supergroup Opisthokonta
Fungi belong to the eukaryotic Supergroup Opisthokonta, along with animals and several groups of unicellular eukaryotes. Opisthokonts are named for cells that, when flagellated, usually have a single posterior flagellum. This feature is not obvious in most modern fungi or animals, but it can still be seen in animal sperm cells and in the motile cells of some early-diverging fungi, especially chytrid fungi. Opisthokonta includes two major branches that are useful for comparison. Holozoa includes animals and their closest relatives, while Holomycota includes fungi and their closest relatives. Both lineages include unicellular forms, and multicellularity evolved independently in the animal and fungal branches. Historically, fungi were classified with plants. Carl Linnaeus originally divided life into two major kingdoms: plants and animals. Because fungi appeared to grow from the ground and lacked obvious movement, they were placed with plants. Later, as biologists learned more about cell structure, nutrition, and reproduction, fungi were separated into their own kingdom. DNA evidence later showed that fungi and animals are both opisthokonts and are more closely related to each other than either is to plants.
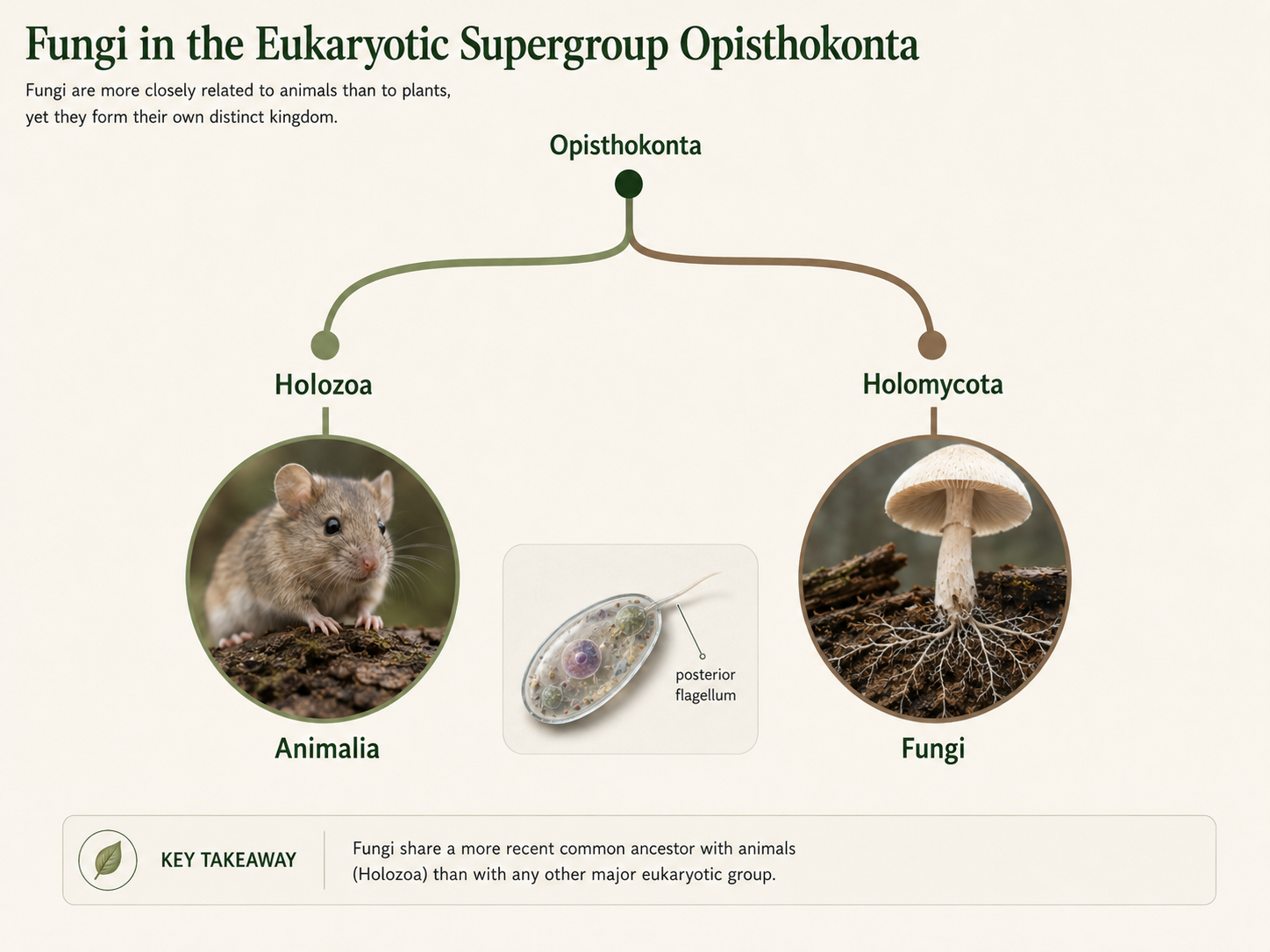
Figure 2. Fungi within Opisthokonta. Fungi and animals are both opisthokonts, but they belong to different evolutionary branches. Animals are part of Holozoa and generally digest food internally, while fungi are part of Holomycota and digest food externally before absorbing nutrients.
Holomycota: External Digestion and Absorptive Feeding
Domain Eukarya: Opisthokonta: Holomycota
Holomycota includes fungi and their closest non-fungal relatives. The closest living relatives of fungi are nucleariids, amoeba-like unicellular organisms that move and feed using thin, thread-like extensions called filose pseudopodia. Nucleariids do not have the chitin-rich cell wall that defines fungi, but their relationship to fungi helps biologists infer what early fungal relatives may have been like. The key shift in fungal evolution was the origin of a body plan built around absorption. Instead of chasing, ingesting, and internally digesting food, fungi evolved external digestion. Hyphae grow into a food source, release enzymes, break large molecules into smaller molecules, and absorb those nutrients across the cell surface. This strategy made fungi especially effective decomposers, parasites, and mutualistic partners. This close relationship between fungi and animals also helps explain why fungal infections can be difficult to treat. Bacteria are prokaryotes, so many antibiotics target structures or pathways that human cells do not have. Fungi, like humans, are eukaryotes, which makes it harder to kill fungal cells without also harming animal cells. Common fungal infections such as athlete’s foot are usually not life-threatening, but they can be persistent because fungi are biologically more similar to us than bacteria are.
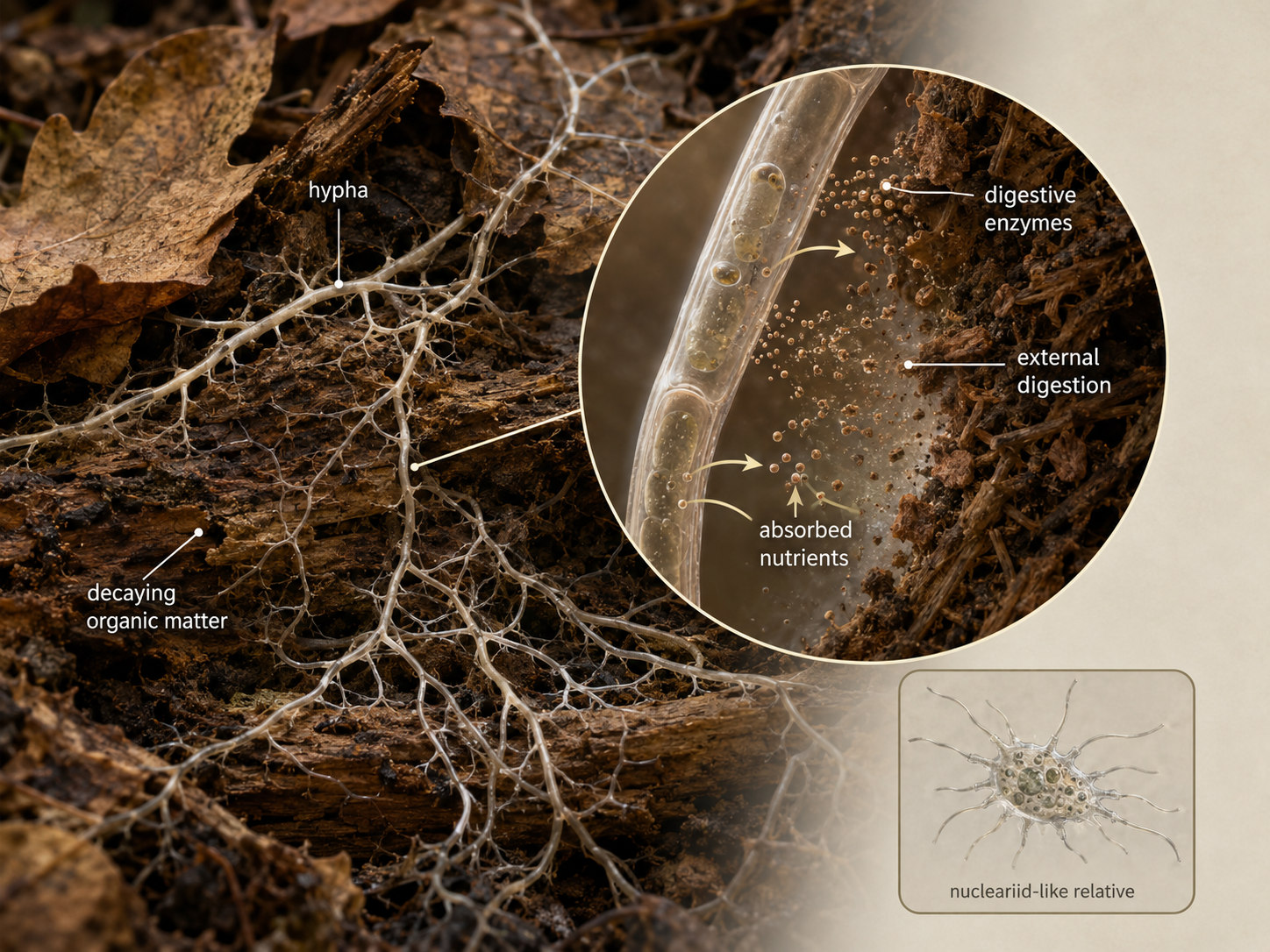
Figure 3. Holomycota and the Origin of Fungal Feeding. Fungi evolved from opisthokont ancestors related to nucleariid-like organisms. The fungal lineage became specialized for external digestion, using chitin-supported cells and absorptive surfaces to obtain nutrients from the environment.
Hyphae, Mycelium, and the Fungal Body Plan
Kingdom Fungi
Hyphae are narrow filaments that can branch repeatedly through a food source, giving fungi a large surface area for releasing enzymes and absorbing nutrients. A dense network of hyphae is called a mycelium, and it can spread through soil, leaf litter, wood, living tissue, or plant roots. This body plan is highly efficient because a fungus does not need to swallow food. Instead, it grows into or across the material it is using as food. In decomposer fungi, hyphae penetrate dead leaves, wood, or animal remains. In parasitic fungi, hyphae invade living hosts. In mutualistic fungi, hyphae grow in association with other organisms, such as plant roots or photosynthetic cells. Hyphae also allow fungi to respond to their environment. When a growing hypha encounters a useful food source, the fungus can direct more growth toward that resource. When a food source is exhausted, parts of the mycelium may die back while the fungus redirects energy toward new areas. Many fungal hyphae are divided into cells by cross-walls called septa. Septa often contain pores that allow cytoplasm, nutrients, organelles, and chemical signals to move between compartments. This internal sharing helps make a multicellular fungus an integrated organism rather than just a loose colony of separate cells. Some fungi produce visible reproductive structures. Mushrooms, puffballs, shelf fungi, and morels are not the entire fungus; they are fruiting bodies that release spores. Feeding hyphae are usually spread out through the food source, while reproductive hyphae are densely packed to form upright structures that help spores disperse. Fungi have an extremely high surface-area-to-volume ratio, which makes them excellent at absorption but also creates a tradeoff: thin hyphae can dry out quickly. Spores help solve this problem because they often have resistant outer walls that allow them to survive dry conditions for long periods before germinating when moisture returns.

Figure 4. Hyphae and Mycelium. Multicellular fungi grow as branching hyphae that form a mycelium. Septa with pores can connect hyphal compartments, allowing materials and signals to move through the fungus. The mycelium increases surface area for external digestion and nutrient absorption, while fruiting bodies such as mushrooms produce and release spores.
Fungal Reproduction and Spores
Kingdom Fungi
Fungi reproduce using spores, small reproductive cells that can disperse through air, water, soil, or animal carriers. Spores allow fungi to spread to new habitats and survive changing environmental conditions. Many fungi reproduce both sexually and asexually, depending on environmental conditions and the fungal group. Asexual reproduction can produce large numbers of genetically similar spores, allowing a fungus to spread quickly when conditions are favorable. Sexual reproduction combines genetic material from compatible mating types and can increase genetic variation. In fungi, “male” and “female” are usually not the best terms because many fungi have mating types rather than sexes, and compatible hyphae or cells can fuse during sexual reproduction. Most fungal spores are non-motile, meaning they do not swim. They are carried passively by wind, water, animals, or physical disturbance. However, chytrid fungi are an important exception because they produce motile cells with flagella. This trait connects chytrids to the opisthokont ancestry of fungi and helps explain why many chytrids live in aquatic or moist environments. In many fungi, sexual reproduction includes unusual nuclear stages. In some groups, especially Ascomycota and Basidiomycota, cells may contain two genetically distinct haploid nuclei for part of the life cycle. This condition is called dikaryotic, and it is a defining feature of the clade Dikarya.

Figure 5. Fungal Spores and Life Cycles. Fungi reproduce with spores. Asexual spores allow rapid dispersal, while sexual reproduction increases genetic variation. Most fungal spores are non-motile and disperse by wind, water, or animals, while chytrid fungi retain motile flagellated cells.
Major Groups of Fungi
Kingdom Fungi
Fungal classification is still being revised as genetic data improves. Some traditional groups are useful for teaching because their members share similar body forms or reproductive structures, but not all of these groups are natural evolutionary clades. A monophyletic group includes an ancestor and all of its descendants. A paraphyletic group leaves out some descendants and therefore does not represent a complete evolutionary branch. For this chapter, the major teaching groups are Microsporidia, Chtridomycota, Zygomycota, Glomeromycota, Ascomycota, and Basidiomycota. Some of these names represent well-supported clades, while others are historically used groupings that should be treated carefully. A simplified teaching phylogeny can be summarized as Opisthokonta → Holomycota → Kingdom Fungi, with major fungal groups including Microsporidia, early-diverging flagellated fungal lineages, zygomycete-type lineages, Glomeromycota, and Dikarya. Dikarya then branches into Ascomycota and Basidiomycota. This tree should be treated as a teaching framework, not a final technical classification, because the deeper branches of fungi, especially chytrid-like and zygomycete-like lineages, remain classification-sensitive.
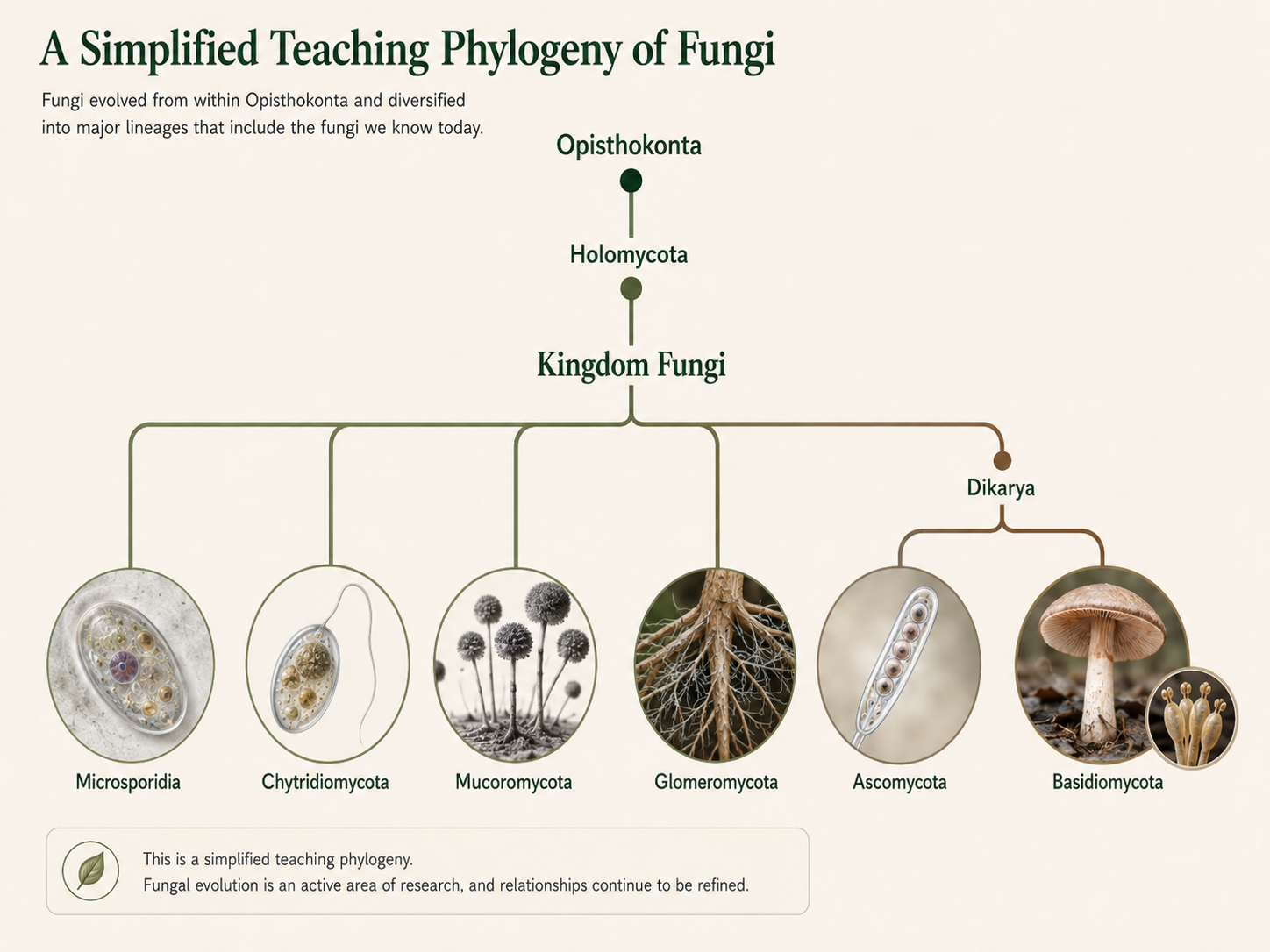
Figure 6. Phylogeny of Fungi. Fungi belong within Opisthokonta and Holomycota. Major fungal groups include Microsporidia, early-diverging flagellated fungal lineages, zygomycete-grade lineages, Glomeromycota, and Dikarya. Dikarya includes Ascomycota and Basidiomycota.
Microsporidia
Kingdom Fungi: Phylum Microsporidia
Microsporidia are unicellular fungi that live as intracellular parasites, mostly inside animal cells. For a long time, they were classified as protists rather than fungi because they are highly reduced and unusual. Genetic evidence now places them within or very close to the fungal lineage. Microsporidia share important fungal traits. They absorb nutrients and possess chitin-containing structures. Their most distinctive feature is the polar tube, a specialized infection structure used to enter host cells. When a microsporidian infects a host, the polar tube can rapidly extend and pierce the host cell membrane, allowing the parasite to deliver its contents into the cell. Once inside, the microsporidian uses the host cell’s resources for growth and reproduction. Microsporidia show that fungi are not limited to mushrooms, molds, and decomposers. Some fungi are microscopic parasites with highly specialized structures for living inside other organisms.
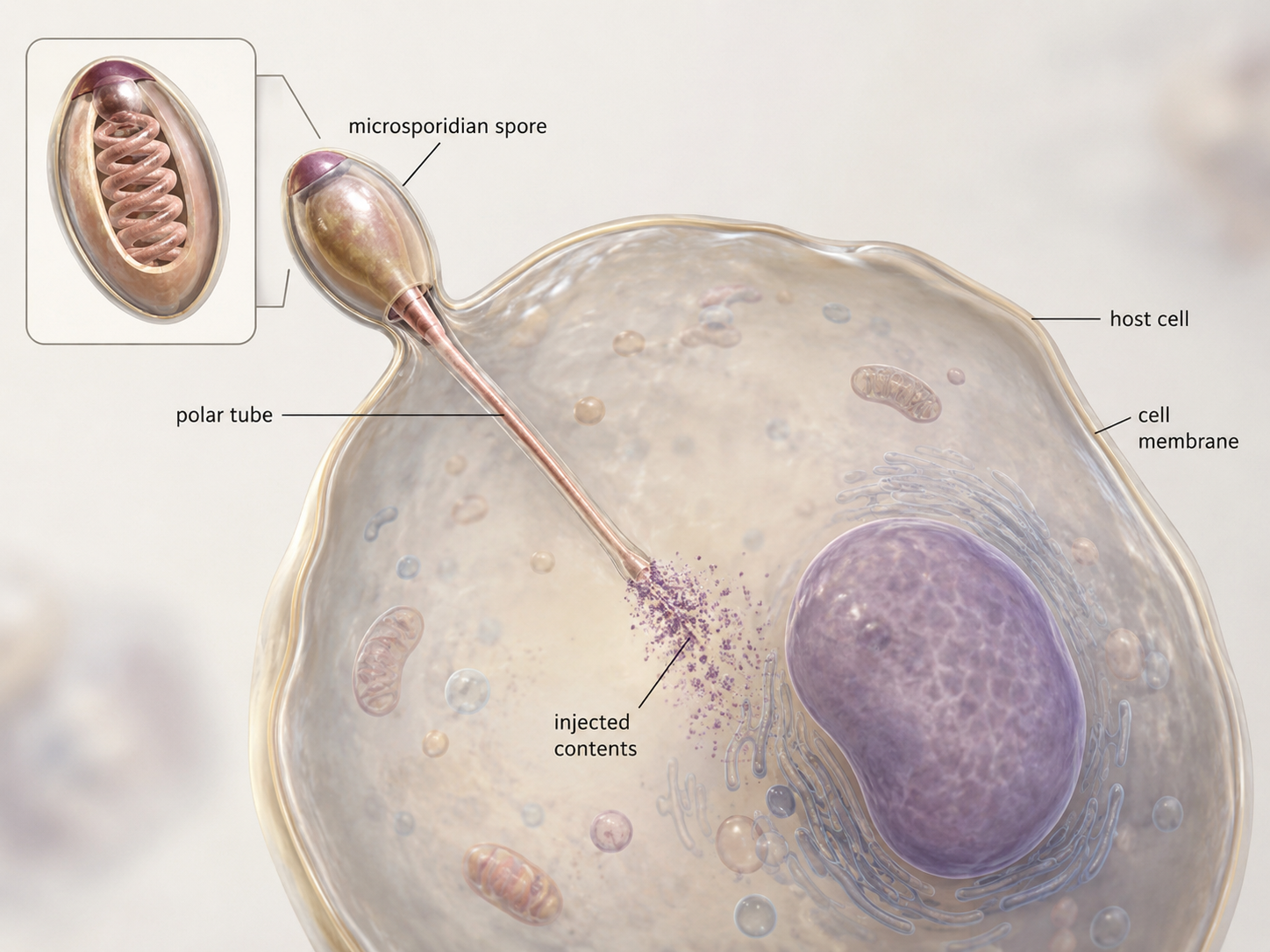
Figure 7. Microsporidia. Microsporidia are unicellular fungal parasites that infect animal cells. They use a specialized polar tube to enter host cells and absorb nutrients from within the host.
Chytrid Fungi: Flagellated Fungal Lineages
Kingdom Fungi: Early-Diverging Flagellated Fungal Lineages; traditionally Phylum Chytridiomycota
Chytrid fungi are mostly aquatic or semi-aquatic fungi. Some are unicellular, while others form simple multicellular structures. Their most important feature is the presence of motile cells with a single flagellum. These motile cells may function as spores or gametes, depending on the life cycle. This flagellated stage is important evolutionarily because most fungi have lost motile cells. Chytrids retain a feature that connects fungi to their opisthokont ancestry. Because swimming cells require water, many chytrids live in ponds, streams, wet soils, or moist tissues. The name Chytridiomycota has often been used for chytrid fungi, but this traditional grouping is complicated. Molecular evidence indicates that flagellated fungi do not all form one simple, neatly bounded group. For our purposes, chytrids are best understood as early-diverging fungal lineages that retain motile cells, rather than as a single settled classification that will never change. Some chytrids are decomposers, breaking down organic material in aquatic ecosystems. Others are parasites. One well-known chytrid-related disease is chytridiomycosis, which has contributed to severe amphibian decline around the world. The disease affects amphibian skin, interfering with water balance, ion regulation, and gas exchange. Because many amphibians rely heavily on their skin for these functions, infection can be deadly.

Figure 8. Chytrid Fungi. Chytrid fungi are mostly aquatic or moisture-dependent fungi that retain motile flagellated cells. This feature links them to early opisthokont traits and distinguishes them from most other fungi, which have non-motile spores.
Zygomycetes: A Historically Used Group of Molds
Kingdom Fungi: historically Phylum Zygomycota
Zygomycetes are a traditional group of mostly terrestrial fungi that include familiar molds such as black bread mold, Rhizopus stolonifer. These fungi commonly grow as hyphae across food, soil, or decaying organic matter. Many reproduce asexually by producing spores inside rounded structures called sporangia. These sporangia are often held upright on specialized hyphae, allowing spores to disperse more easily. During sexual reproduction, compatible mating types can fuse and produce a thick-walled structure called a zygosporangium. This resistant structure can survive difficult environmental conditions and later produce spores when conditions improve. This is especially important in terrestrial environments, where spores and reproductive structures must resist drying out. Zygomycota is now treated carefully because it is not considered a single well-supported monophyletic group in the traditional sense. Instead, fungi once grouped as zygomycetes appear to belong to several related lineages. The term is still useful for describing molds with similar reproductive structures, but realize that it is a simplified and historically used classification, and does not represent modern phylogenetic analysis. The uncertainty surrounding chytrid-like and zygomycete-like fungi is a good example of how classification changes with evidence. Traditional classifications were often based on visible reproductive structures. DNA evidence has shown that similar structures do not always mean all members belong to one natural clade. Some traits may have evolved more than once, or they may have been present in ancestors and later lost in some lineages.
Figure 9. Zygomycetes. Zygomycetes include familiar molds such as black bread mold. They often reproduce asexually using sporangia and can reproduce sexually by forming resistant zygosporangia. The traditional group Zygomycota is historically useful but not a settled monophyletic clade.

Glomeromycota and Arbuscular Mycorrhizae
Kingdom Fungi: Phylum Glomeromycota
Glomeromycetes are fungi best known for forming arbuscular mycorrhizae, mutualistic associations with plant roots. The word mycorrhiza means “fungus-root.” In this relationship, fungal hyphae grow through soil and into close contact with root cells. Inside the root, the fungus forms highly branched structures called arbuscules, which increase the surface area for exchange between the plant and the fungus. This partnership benefits both organisms, known as a mutualism. The fungal hyphae extend far into the soil and absorb water and mineral nutrients more efficiently than roots alone. The plant receives some of these resources from the fungus, and in return, the plant provides sugars produced by photosynthesis. Mycorrhizal fungi are especially important for phosphorus uptake. Phosphorus is a key part of nucleic acids, cell membranes, and ATP. Some plant roots have difficulty accessing phosphate efficiently on their own. Fungal hyphae are much thinner than roots and can explore a larger volume of soil, giving the plant access to phosphorus and other minerals that would otherwise be difficult to obtain. The benefit of mycorrhizae is both physical and chemical. Physically, fungal hyphae increase the absorptive surface area beyond the root system. Chemically, fungi can help mobilize nutrients from soil particles and organic matter. In return, the fungus receives a relatively steady supply of carbohydrates from the plant. Arbuscular mycorrhizae are extremely widespread among land plants and likely played an important role in the early evolution of plants on land. They show that fungi are not only decomposers or pathogens. Many fungi are essential mutualists that help plants survive, grow, and acquire nutrients.
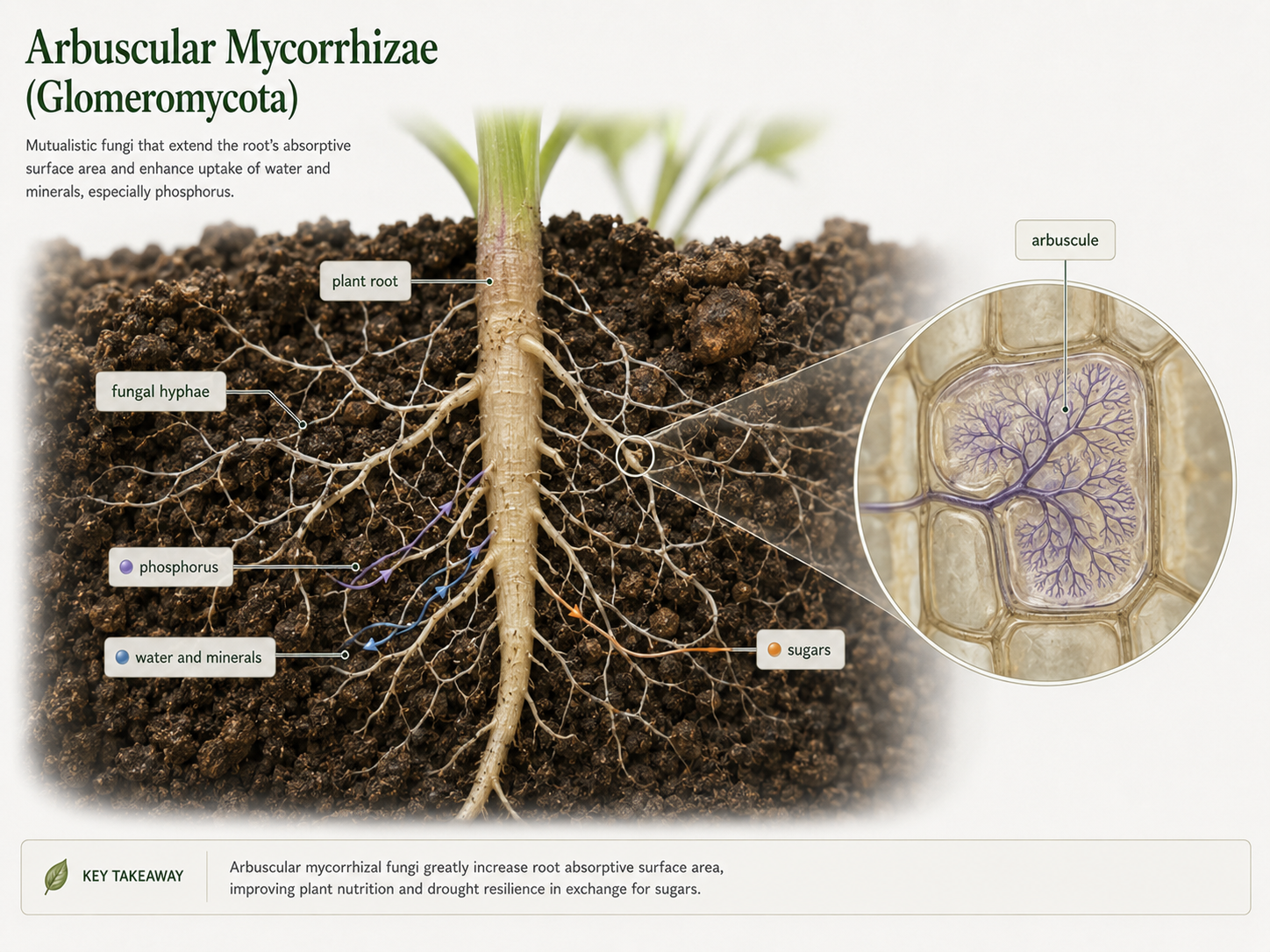
Figure 10. Glomeromycota and Arbuscular Mycorrhizae. Glomeromycetes form mutualistic relationships with plant roots. Fungal hyphae absorb water and minerals from soil, including phosphorus, while plants provide sugars from photosynthesis. Arbuscules inside roots increase surface area for nutrient exchange.
Dikarya: Fungi with a Two-Nucleus Stage
Kingdom Fungi: Subkingdom Dikarya
Dikarya is a major fungal clade that includes Ascomycota and Basidiomycota. These fungi are united by a distinctive life-cycle stage in which cells may contain two genetically distinct haploid nuclei. This condition is called dikaryotic. The dikaryotic stage is unusual because the nuclei share the same cell without immediately fusing. Nuclear fusion occurs later during sexual reproduction. This pattern gives Dikarya a distinctive reproductive biology and helps explain why ascomycetes and basidiomycetes are grouped together. Dikarya includes many of the fungi most familiar to people: mushrooms, puffballs, shelf fungi, morels, truffles, yeasts, molds, rusts, smuts, and many lichens. These fungi play major roles as decomposers, mutualists, pathogens, and food sources. Because both Ascomycota and Basidiomycota have dikaryotic stages, biologists infer that the common ancestor of these two groups also likely had this trait. This is an example of how shared traits can be used to infer features of ancestral organisms.
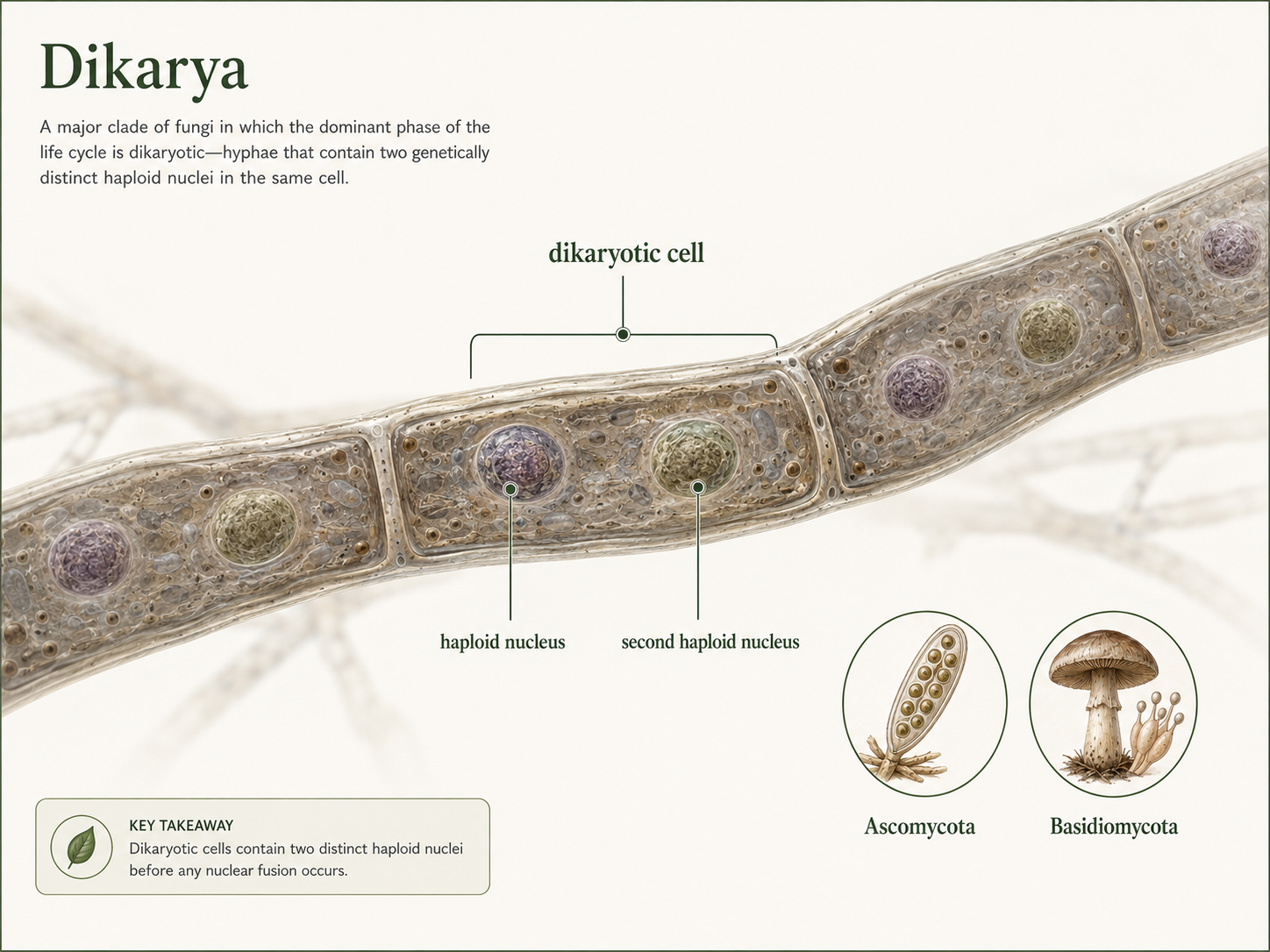
Figure 11. Dikarya. Dikarya includes Ascomycota and Basidiomycota. Members of this clade can have a dikaryotic stage, in which cells contain two genetically distinct haploid nuclei before nuclear fusion occurs later in reproduction.
Ascomycota: Sac Fungi
Kingdom Fungi: Subkingdom Dikarya: Phylum Ascomycota
Ascomycota are known as sac fungi because they produce sexual spores inside sac-like structures called asci. A single ascus often contains eight spores, although the number can vary. These asci may be grouped together in larger reproductive structures called ascocarps. Some ascocarps are cup-shaped, such as apothecia, while others take different forms. Ascomycota is one of the largest and most diverse fungal groups. It includes yeasts, many molds, morels, truffles, cup fungi, plant pathogens, and many lichen-forming fungi. Some ascomycetes are unicellular, while others are multicellular. Some are decomposers, others are parasites, and many form mutualistic relationships. Ascomycetes are important to humans and ecosystems. Yeasts are used in bread, beer, wine, and biotechnology. Brewer’s yeast produces carbon dioxide during fermentation, which causes bread dough to rise. Yeast fermentation also converts sugars into ethanol in beer and wine production. Morels and truffles are valued edible fungi. Other ascomycetes cause plant diseases or produce medically important compounds. Their diversity shows how one fungal body plan can be modified for many ecological roles.

Figure 12. Ascomycota. Ascomycetes are sac fungi that produce sexual spores inside asci. This group includes yeasts, molds, morels, truffles, cup fungi, many lichen-forming fungi, and important decomposers, pathogens, and mutualists.
Lichens: Fungal Partnerships with Photosynthetic Cells
Mostly Kingdom Fungi: Phylum Ascomycota, with some Basidiomycota
Lichens are symbiotic associations between a fungus and a photosynthetic partner, usually a green alga or cyanobacterium. The fungal partner forms most of the visible body, called the thallus. It provides structure, protection, anchorage, and access to water and minerals. The photosynthetic partner produces sugars through photosynthesis and shares some of that carbon with the fungus. Most lichens involve ascomycete fungi, although some involve basidiomycetes. A lichen is not a single organism in the same sense as a mushroom or a plant. It is a stable partnership that functions as an ecological unit. Lichens can survive in harsh environments, including bare rock, tree bark, deserts, tundra, and exposed surfaces with little soil. They can contribute to soil formation by slowly weathering rock, trapping particles, and adding organic material after they die. Lichens also provide food and habitat for small organisms and can be sensitive indicators of air quality.

Figure 13. Lichens. A lichen is a symbiotic partnership between a fungus and a photosynthetic alga or cyanobacterium. The fungus forms the thallus and provides protection and structure, while the photosynthetic partner provides carbohydrates.
Basidiomycota: Club Fungi
Kingdom Fungi: Subkingdom Dikarya: Phylum Basidiomycota
Basidiomycota are known as club fungi because they produce sexual spores on club-shaped structures called basidia. The word basidium refers to a small pedestal-like structure. Spores called basidiospores are often produced in groups of four on each basidium, although variation occurs. The visible mushroom is a reproductive structure that elevates basidia so spores can disperse. Basidiomycota includes many familiar fungi, including mushrooms, puffballs, shelf fungi, chanterelles, stinkhorns, rusts, and smuts. Most basidiomycetes are multicellular, although some yeast-like forms exist. Many grow as extensive networks of hyphae within soil, wood, or living hosts. Basidiomycetes are especially important decomposers. Many species can break down wood, including lignin, a complex structural polymer that helps make plant cell walls tough and resistant to decay. Wood-decay fungi play a major ecological role because they release carbon and nutrients locked inside dead trees and woody debris. Without fungi, forests would accumulate far more undecomposed plant material. Not all basidiomycetes are decomposers. Rusts and smuts are plant pathogens that can infect crops and wild plants. Corn smut is one example of a fungal disease that can damage agricultural crops. Others form mycorrhizal relationships with trees, helping plants absorb nutrients while receiving sugars in return.

Figure 14. Basidiomycota. Basidiomycetes are club fungi that produce basidiospores on basidia. This group includes mushrooms, puffballs, shelf fungi, rusts, smuts, and many major wood-decay fungi.
Fungi as Decomposers
Kingdom Fungi
Fungi are among the most important decomposers in terrestrial ecosystems. They break down dead organisms, leaf litter, wood, and organic waste. This releases nutrients back into ecosystems and makes those nutrients available to plants, microbes, and other organisms. Without decomposers, forests would be buried under massive accumulations of dead plant material. Fallen trees, leaves, roots, and animal remains would pile up instead of being recycled. Fungi help prevent that buildup by breaking down complex organic materials and returning carbon, nitrogen, phosphorus, and other elements to the living world. Fungi are especially important because they can break down materials that many organisms cannot digest easily. Plant cell walls contain cellulose, a tough polysaccharide. Wood also contains lignin, a complex structural polymer that makes woody tissue rigid and resistant to decay. Many fungi are highly effective at breaking down cellulose, and some fungi, especially many basidiomycetes, are major decomposers of lignin-rich wood. This decomposition makes fungi central to nutrient cycling. The dead become resources for the living.

Figure 15. Fungi as Decomposers. Fungi break down dead organisms, leaf litter, cellulose-rich plant material, and lignin-rich wood. This releases nutrients and carbon compounds back into ecosystems.
Fungi and the Carbon Cycle
Kingdom Fungi
The carbon cycle describes how carbon moves through ecosystems and the atmosphere. Plants and other photosynthetic organisms capture carbon dioxide from the air and use it to build sugars and other organic molecules. This carbon becomes part of leaves, wood, roots, fruits, seeds, and other living tissues. Animals, fungi, and many microbes then use this organic carbon for food. Fungi return carbon to the cycle through decomposition and cellular respiration. As fungi break down plant material, they absorb organic molecules and use them for energy and growth. During cellular respiration, fungi release carbon dioxide back into the atmosphere. That carbon dioxide can then be taken up again by photosynthetic organisms. Fungi therefore connect plant growth, decomposition, soil carbon, and atmospheric carbon dioxide. Their activity helps determine how quickly carbon moves from dead organic matter back into the atmosphere or into soil ecosystems.
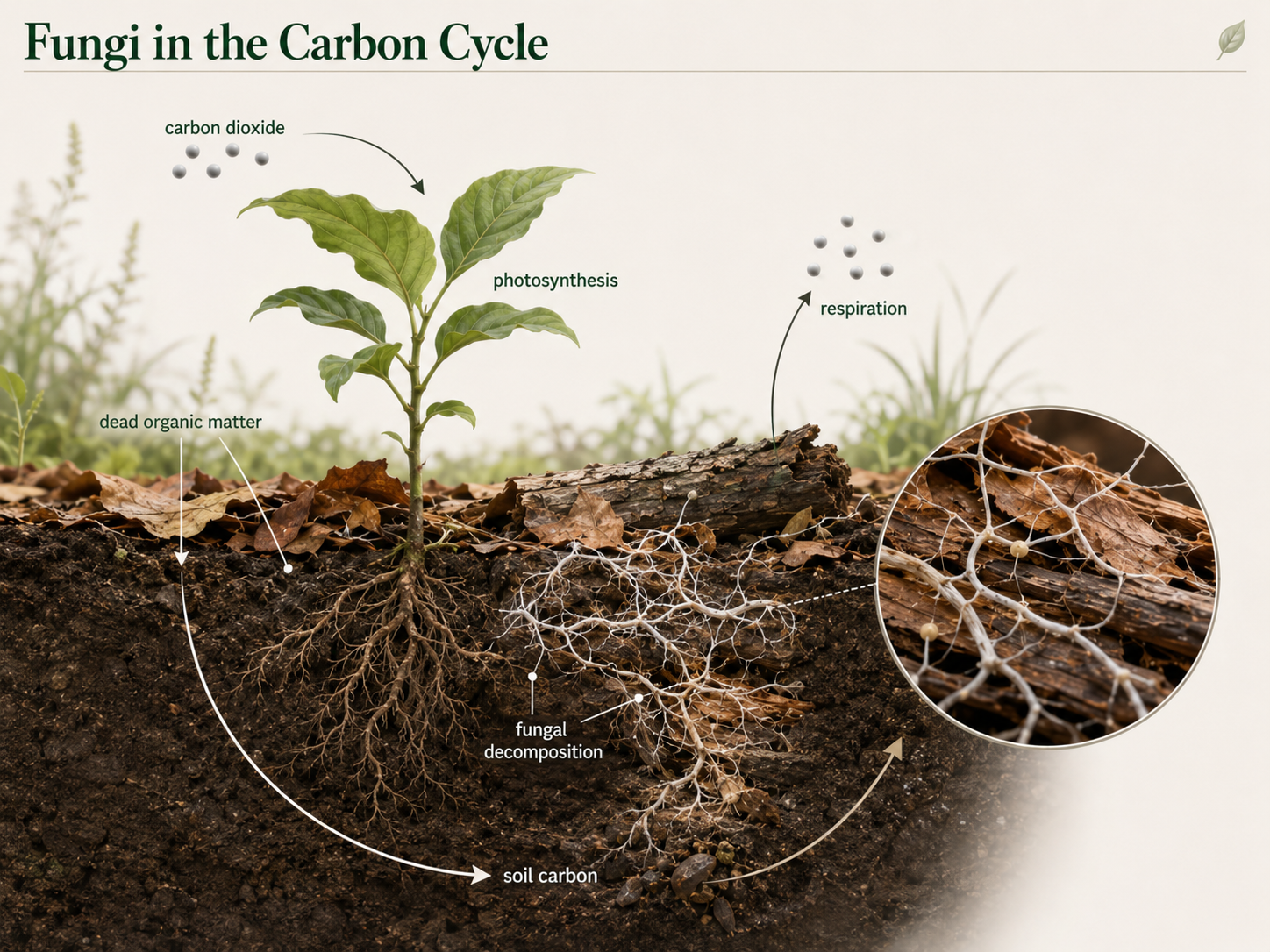
Figure 16. Fungi and the Carbon Cycle. Photosynthetic organisms capture carbon dioxide and store carbon in organic molecules. Fungi decompose dead organic material, use some carbon for growth, and release carbon dioxide through cellular respiration.
Fungi and Human Society
Kingdom Fungi
Fungi are deeply important to human life. Some fungi are eaten directly as mushrooms, including cultivated varieties such as portobello, cremini, shiitake, and button mushrooms. Other edible fungi, such as morels and truffles, are collected from natural habitats and are highly valued as food. Fungi are also essential in fermentation. Yeasts convert sugars into carbon dioxide and ethanol. In bread, carbon dioxide bubbles cause dough to rise. In beer and wine, yeast fermentation produces alcohol. Fungi also contribute to the production of some cheeses and other fermented foods. More broadly, fermentation has been important in human food storage, cuisine, and agriculture, especially before refrigeration. Fungi have also transformed medicine. Penicillin, the first widely used antibiotic, was discovered from a fungus in the genus Penicillium. Antibiotic production gives some fungi an advantage in competition with bacteria, because fungi and bacteria often compete for the same resources. Humans have used this fungal chemical warfare as medicine. Fungi can also cause economic damage. Fungal diseases affect crops, stored foods, forests, and buildings. They can be harder to control than bacterial diseases because fungi are eukaryotes, making it more difficult to target them without harming plants, animals, or humans.

Figure 17. Fungi and Human Society. Fungi provide food, fermentation products, antibiotics, and biotechnology tools, but they also cause crop diseases, food spoilage, and human infections.