
Chapter: Community Ecology
A community is made up of all the populations of all the species living and interacting in a given area. A forest community, for example, includes trees, shrubs, fungi, insects, birds, mammals, bacteria, and many other organisms. A pond community includes algae, aquatic plants, fish, insects, amphibians, snails, bacteria, and decomposers. The key point is that a community is not just one species. It is a living network of many species interacting in the same place. Community ecology is the study of how species interact with one another and how those interactions shape the structure, diversity, and change of ecological communities over time. Community ecologists ask questions such as: Why do some species live together while others do not? Why are some communities more diverse than others? How do predators affect prey populations? How does competition shape the traits of species? Why do some communities recover quickly after disturbance while others change dramatically? These questions matter because species do not live in isolation. Every organism is affected by competitors, consumers, mutualists, parasites, pathogens, decomposers, and the physical environment. Community ecology connects population ecology to ecosystem ecology. Population ecology focuses on how one species changes in abundance. Community ecology asks how multiple species influence one another. Ecosystem ecology then adds energy flow and nutrient cycling. Together, these fields help explain how the living world is organized.
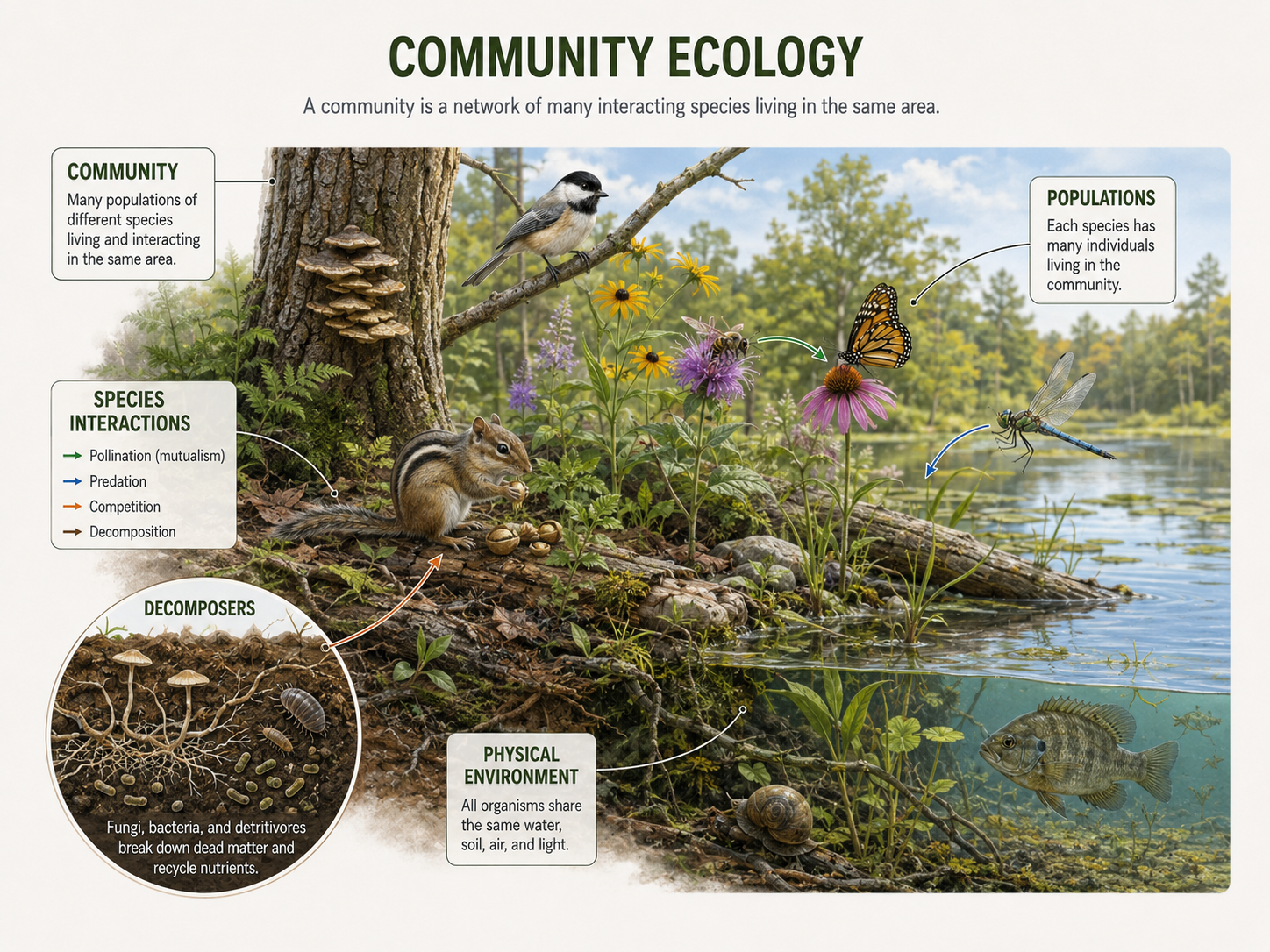
Figure 1. Community Ecology. A community includes all populations of all species living and interacting in a given area. Community ecology studies how species interactions shape the structure, diversity, and change of ecological communities over time.
Species Interactions
Species interact in several major ways, and these interactions can be described by how they affect the fitness, survival, or reproduction of each species involved. Ecologists often summarize these effects using plus, minus, and zero symbols. A plus sign means a species benefits. A minus sign means a species is harmed. A zero means the species is not strongly affected. Competition occurs when two organisms use the same limited resource, such as food, space, light, water, nesting sites, minerals, or nutrients. Because both organisms have reduced access to the resource, competition is a negative-negative interaction (-/-). Consumption occurs when one organism feeds on another organism. This includes predation, herbivory, and parasitism. The consumer benefits, while the organism being consumed is harmed, making consumption a positive-negative interaction (+/-). Mutualism occurs when both species benefit from the interaction. Pollination is a familiar example: flowering plants gain help with reproduction, while pollinators gain nectar, pollen, or other food rewards. Mutualism is a positive-positive interaction (+/+). Commensalism occurs when one species benefits and the other is not significantly helped or harmed. This is a positive-neutral interaction (+/0). True commensalism can be difficult to prove because subtle costs or benefits may be hard to measure. Amensalism occurs when one species is harmed while the other is not affected. This is a negative-neutral interaction (-/0). For example, a large tree may shade out small plants below it without being affected by those plants. These categories are useful, but real species interactions can be more complicated. The same interaction may change depending on environmental conditions, population density, life stage, or season.

Figure 2. Species Interactions. Species interactions can be described by how they affect each species involved. Competition harms both species, consumption benefits one and harms the other, mutualism benefits both, commensalism benefits one without strongly affecting the other, and amensalism harms one without strongly affecting the other.
Competition
Competition is a negative-negative interaction (-/-) because both organisms experience reduced access to a limited resource. A resource is something an organism needs and uses, such as food, light, water, shelter, nesting sites, mates, or space. Competition occurs only when the resource is limited. If there is enough of a resource for everyone, competition is weak or absent. Intraspecific competition occurs among individuals of the same species. For example, young pine trees growing close together compete for light, water, and soil nutrients. Deer in the same population may compete for food during winter. Intraspecific competition often becomes stronger as population density increases. Interspecific competition occurs between individuals of different species. For example, two bird species may compete for similar nesting cavities, or two plant species may compete for sunlight in the same forest gap. Competition can be direct or indirect. In direct competition, organisms physically interfere with one another. A bird may defend a territory from another bird, or a plant may release chemicals into the soil that inhibit nearby plants. In indirect competition, organisms do not physically interact, but they reduce a shared resource. Two species of insects feeding on the same plant may never meet, but each reduces the food available to the other. Competition matters because it can influence population size, species distributions, behavior, and evolution. Over time, competition can favor traits that reduce overlap between species, allowing them to coexist by using resources in different ways.
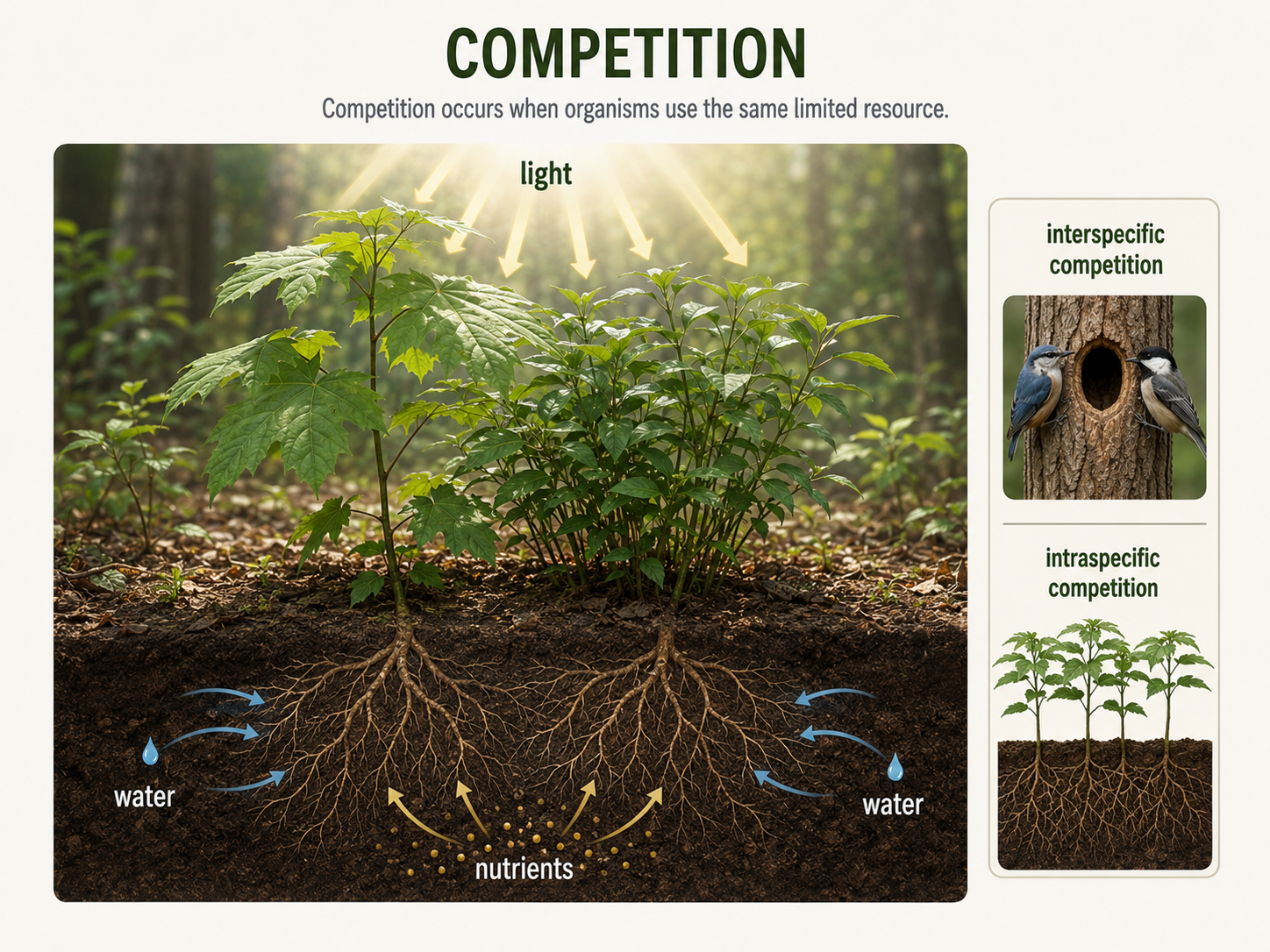
Figure 3. Competition. Competition occurs when organisms use the same limited resource. It can occur within a species or between species and can shape population size, behavior, species distributions, and evolution.
Niches and Resource Use
A species’ niche describes how it fits into its environment. A niche includes the conditions a species can tolerate, the resources it uses, and the role it plays in the community. Temperature, moisture, food type, nesting location, predators, competitors, parasites, mutualists, and timing of activity can all be part of a species’ niche. A niche is not just what a species does. It also includes where it can live, what it can tolerate, what it eats, what eats it, when it is active, how it reproduces, and how it interacts with other species. The niche model predicts that species are more likely to coexist when they use resources differently. When species reduce overlap in resource use, competition becomes weaker in the niche space they overlap. This is called resource partitioning. Darwin’s finches are a classic example. On the Galápagos Islands, different finch species often have different beak sizes and shapes. These differences allow them to specialize on different food sources, such as small seeds, large seeds, or insects. Resource partitioning can occur in several ways. Species may eat different foods, feed in different places, feed at different times, nest in different habitats, or tolerate different physical conditions. For example, different warbler species may feed in different parts of the same tree. Some search for insects near the top, others near the middle, and others near the trunk. Over evolutionary time, competition can also lead to character displacement, where similar species become more different when they live together than when they live apart.

Figure 4. Niches and Resource Use. A species’ niche includes the conditions it tolerates, the resources it uses, and its role in the community. Resource partitioning can reduce competition by allowing similar species to use different foods, places, times, or conditions.
Fundamental and Realized Niches
A species’ fundamental niche is the full range of environmental conditions and resources it could use if there were no competitors, predators, parasites, or other limiting species interactions. For example, a plant species may be physiologically capable of living across a wide range of soil moisture levels. A species’ realized niche is the actual range of conditions and resources it uses in nature after biotic interactions are included. Competitors, predators, herbivores, parasites, pathogens, and mutualists can all change where a species actually lives and how it uses resources. For example, a plant may be able to tolerate both moderately dry and moderately wet soils. However, if a stronger competitor dominates the wetter part of that range, the weaker competitor may only persist in drier areas. In that case, the plant’s realized niche is smaller than its fundamental niche. A classic example involves barnacles living along rocky shorelines. One barnacle species may be able to survive lower on the shore, but it may be excluded from that zone by a stronger competitor. Another species may be restricted to higher areas because it cannot tolerate being submerged for long periods. The realized distribution of each species reflects both physical conditions and species interactions. This distinction is important because species are not limited only by climate or physical conditions. They are also limited by the other organisms around them.

Figure 5. Fundamental and Realized Niches. A fundamental niche is the full range of conditions and resources a species could use without limiting species interactions. A realized niche is the range actually used in nature after competition, predation, parasitism, mutualism, and other interactions are included.
Competitive Exclusion Principle
The competitive exclusion principle states that two species cannot coexist indefinitely if they require exactly the same limiting resources in the same environment. In simple terms, complete competitors cannot coexist. This principle is often associated with G. F. Gause, who studied competition in the protists Paramecium aurelia and Paramecium caudatum. When grown separately, both species increased in population size. When grown together under the same laboratory conditions, one species outcompeted the other, and the weaker competitor declined. Competitive exclusion does not mean that similar species can never live in the same community. Instead, it means that coexistence usually requires some difference in niche, resource use, behavior, habitat, timing, or environmental tolerance. Species may coexist by feeding at different times, using different parts of a habitat, specializing on different food sizes, or tolerating different environmental conditions. Competitive exclusion is easiest to observe in simple laboratory systems, but nature is more complex. Environmental variation, disturbance, predation, mutualism, migration, and chance events can all prevent one competitor from completely eliminating another. Still, the principle is important because it explains why niche differences matter. If two species are too similar in their resource needs, one is expected to become rare, shift its niche, or disappear from that local community.
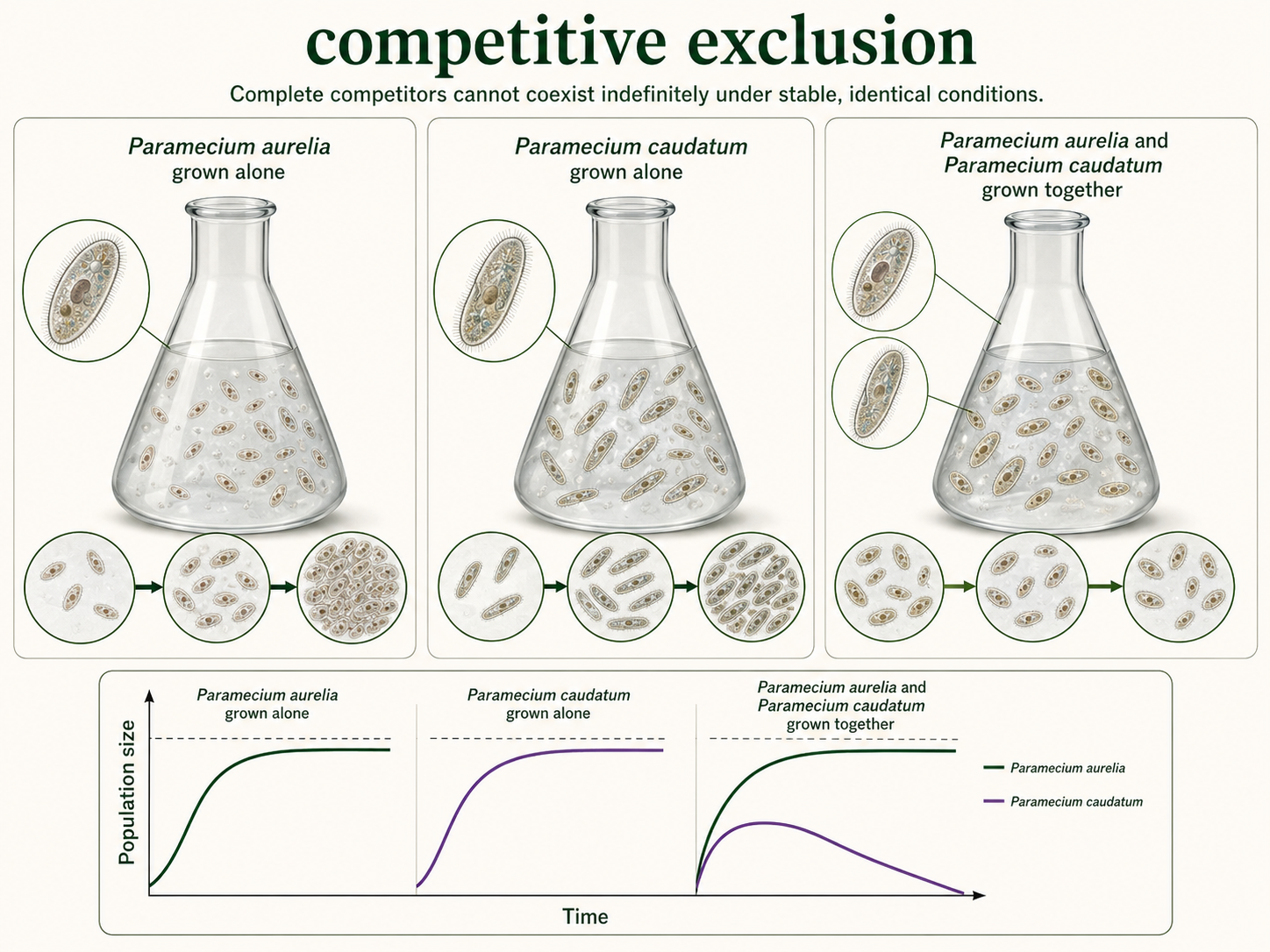
Figure 6. Competitive Exclusion. The competitive exclusion principle states that two species cannot coexist indefinitely if they require exactly the same limiting resources in the same environment. Gause’s Paramecium experiments helped demonstrate this pattern in simple laboratory systems.
Symmetric and Asymmetric Competition
Competition can be symmetric or asymmetric. In symmetric competition, both species are affected in roughly similar ways. Each species reduces the fitness of the other by a comparable amount. In asymmetric competition, one species is affected much more strongly than the other. A stronger competitor may greatly reduce the growth, survival, or reproduction of a weaker competitor while experiencing only a small cost itself. Plant competition is often asymmetric when light is the limiting resource. A taller plant may shade a shorter plant and greatly reduce its photosynthesis. The shorter plant may have little effect on the taller plant. In this case, the competition is strongly unequal. Most real examples fall somewhere between these two extremes. Competition is often a continuum rather than a simple category. Understanding the strength and symmetry of competition helps ecologists predict which species are likely to coexist and which species may exclude others.
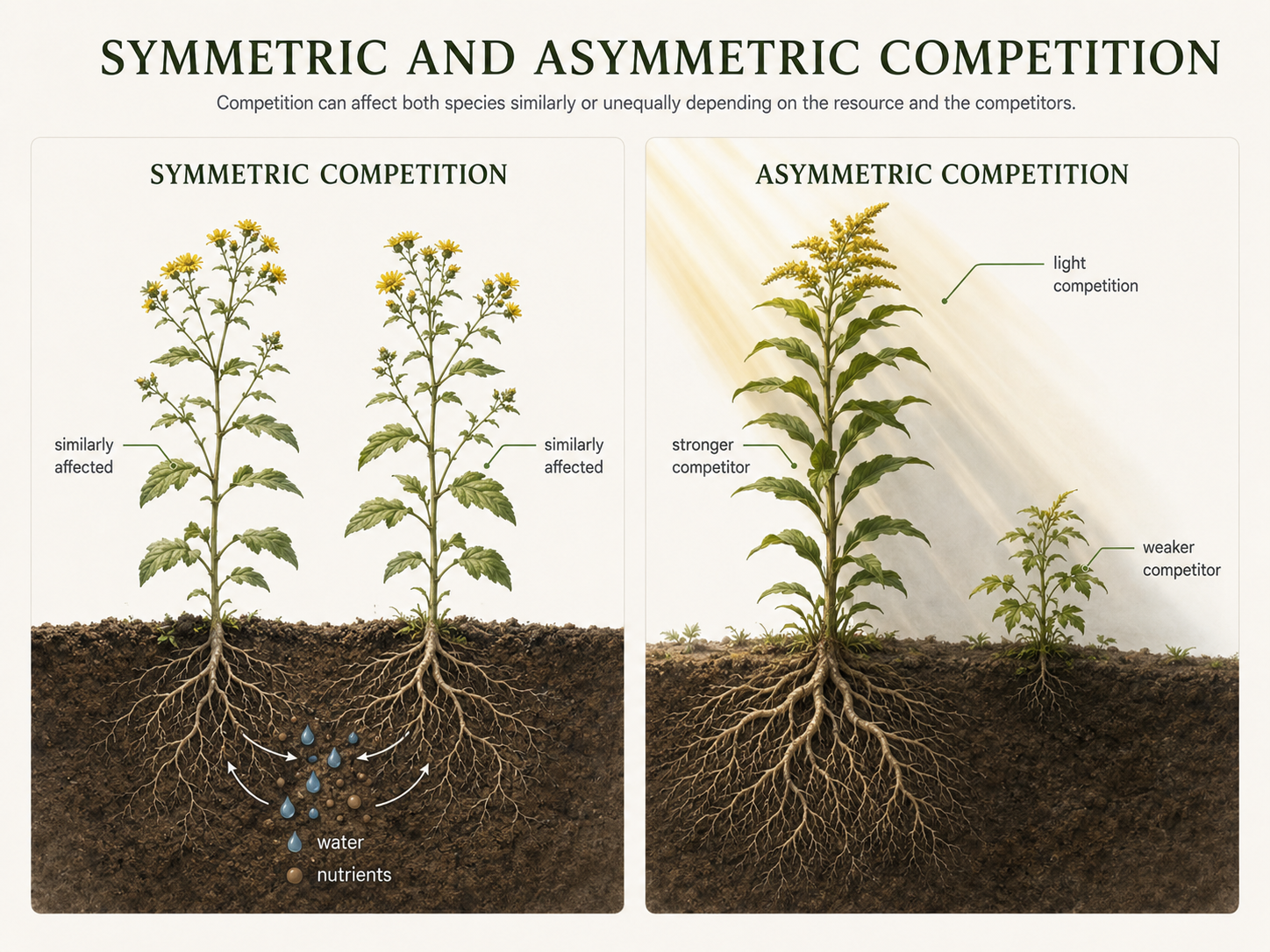
Figure 7. Symmetric and Asymmetric Competition. Competition is symmetric when both species are affected in roughly similar ways. Competition is asymmetric when one species is harmed much more strongly than the other, such as when a tall plant shades a shorter plant.
Consumption
Consumption occurs when one organism obtains energy and nutrients by feeding on another organism or on part of another organism. Predation occurs when one animal kills and eats another animal. A hawk eating a mouse, a spider eating a fly, or a shark eating a fish are examples of predation. Herbivory occurs when an animal eats plant tissue. A caterpillar eating leaves, a deer browsing shrubs, or a grasshopper feeding on grass are examples of herbivory. Herbivory usually does not kill the plant immediately, but it can reduce growth, reproduction, and survival. Parasitism occurs when one organism lives on or in a host and consumes resources from that host without usually killing it immediately. Ticks, fleas, tapeworms, mistletoe, and many disease-causing organisms are parasites. Parasitoids are a special category, common among some wasps and flies. A parasitoid lays eggs on or inside a host. The developing young feed on the host and eventually kill it. Parasitoids are important in insect communities and are sometimes used in biological pest control. Consumption is a major force in community ecology because it affects population size, behavior, defenses, and evolution. Consumers can reduce prey or host populations, but prey and hosts also evolve traits that make them harder to find, capture, eat, or digest.

Figure 8. Consumption. Consumption occurs when one organism obtains energy and nutrients by feeding on another organism or part of another organism. Predation, herbivory, parasitism, and parasitoid interactions are all forms of consumption.
Coevolutionary Arms Races
A coevolutionary arms race occurs when two interacting species place reciprocal selective pressure on each other. One species evolves a trait, and the other species evolves a response. Predators may evolve better speed, sharper senses, stronger jaws, venom, camouflage, or hunting strategies. Prey may evolve better escape behavior, armor, toxins, warning colors, camouflage, spines, shells, or group defense. The key idea is not that evolution has a goal. Instead, natural selection favors traits that improve survival and reproduction in a particular ecological interaction. As one species changes, it can alter the selective pressures acting on the other. Plants and herbivorous insects provide many examples. A plant may evolve chemical compounds that make its leaves toxic or hard to digest. Some insects may then evolve enzymes that allow them to tolerate those chemicals. In response, plants may evolve new or stronger defenses. Over long periods of time, this back-and-forth selection can produce highly specialized relationships. Parasites and hosts also show coevolution. Hosts evolve immune defenses, while parasites evolve ways to avoid or overcome those defenses. This helps explain why diseases can change rapidly and why host immune systems are under strong evolutionary pressure.
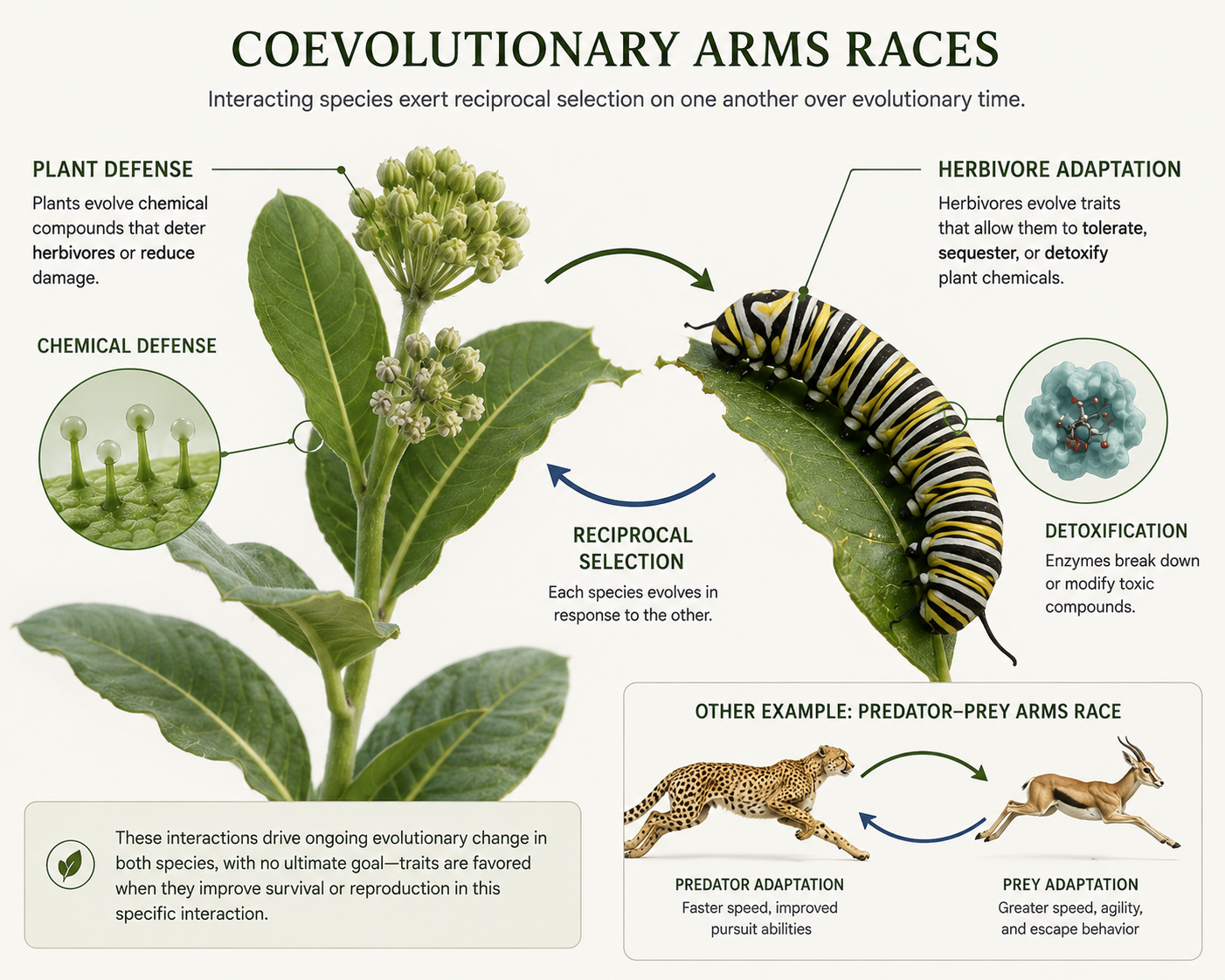
Figure 9. Coevolutionary Arms Races. A coevolutionary arms race occurs when two interacting species place reciprocal selective pressure on each other. As one species evolves a defensive or offensive trait, the other may evolve a response.
Defenses Against Consumption
Because consumption is common, many organisms have evolved defenses against predators, herbivores, and parasites. Some species use avoidance, in which they hide, flee, become active at night, live in hard-to-reach places, or use camouflage to reduce detection. Camouflage, also called cryptic coloration, allows an organism to blend into its surroundings. A walking stick insect resembles a twig. A flounder blends into the ocean floor. Many moths resemble bark or dead leaves. Some species use physical defenses, such as shells, thorns, spines, armor, tough leaves, bark, or tusks. Porcupine quills, cactus spines, turtle shells, and the hard outer coverings of beetles are examples of physical defenses. Some species use chemical defenses. Poison dart frogs, monarch butterflies, milkweed plants, and many fungi produce or store toxic compounds that discourage consumers. Some plants produce bitter chemicals, sticky resins, or compounds that interfere with digestion. Some species use warning coloration, also called aposematic coloration. Bright colors can signal that an organism is toxic, venomous, distasteful, or dangerous. These signals help predators learn to avoid the organism, reducing the chance that it will be attacked in the future. Some animals use group defense. Fish schools, bird flocks, and mammal herds can reduce the chance that any one individual is captured. Groups may confuse predators, increase vigilance, or allow individuals to defend one another. Defenses are costly, so organisms usually face trade-offs. Energy used to produce toxins, armor, or spines cannot also be used for growth or reproduction. Natural selection favors defenses when their survival benefits outweigh their costs.

Figure 10. Defenses Against Consumption. Organisms use many defenses against predators, herbivores, and parasites, including camouflage, physical defenses, chemical defenses, warning coloration, and group defense. These defenses can improve survival but often involve trade-offs.
Mimicry
Mimicry occurs when one species evolves to resemble another species or object. Mimicry can reduce predation, improve hunting success, or increase reproductive success. In Batesian mimicry, a harmless or edible species resembles a harmful, toxic, or dangerous species. For example, some harmless flies resemble wasps or bees. Predators that avoid wasps may also avoid the harmless mimics. In Müllerian mimicry, two or more harmful or unpalatable species resemble one another. This benefits all of them because predators learn more quickly to avoid that shared warning pattern. Many toxic butterflies share similar color patterns for this reason. Mimicry can also be aggressive. In aggressive mimicry, a predator or parasite resembles something harmless or attractive in order to approach prey. Some anglerfish use a lure that resembles prey food. Some snapping turtles use a tongue-like lure that resembles a worm. Prey approach the lure and are captured. Mimicry works because organisms use signals to make decisions. Predators, prey, pollinators, and mates all respond to visual, chemical, auditory, or behavioral cues. When one species evolves to imitate those cues, it can gain a survival or reproductive advantage. Some plants use mimicry to attract pollinators. In pseudocopulation, a flower resembles a female insect so closely that a male insect attempts to mate with it. During this process, the insect may pick up or deposit pollen. Some orchids are famous for this strategy. They may mimic the shape, color, texture, and chemical signals of female bees or wasps. In some cases, they release compounds similar to insect sex pheromones. The male insect is attracted to the flower, attempts to mate, and unintentionally helps pollinate the plant. This is an example of how natural selection can shape highly specialized interactions between plants and animals. It also shows that pollination is not always a simple exchange of food for service.

Figure 11. Mimicry. Mimicry occurs when one species evolves to resemble another species or object. Mimicry can reduce predation, improve hunting success, or increase reproductive success by taking advantage of signals used by other organisms.
What Controls Herbivores?
Herbivore populations can be controlled by both predators and food supply. Top-down control occurs when predators, parasites, or disease limit herbivore populations. For example, wolves may reduce deer numbers or change where deer feed. In this case, consumers at higher trophic levels influence organisms below them. Bottom-up control occurs when plant abundance, plant quality, nutrients, or primary productivity limit herbivore populations. If plants are scarce or low in nutrients, herbivore populations may remain small even if predators are rare. In real ecosystems, herbivores are usually controlled by both top-down and bottom-up forces. Predators, food availability, plant defenses, climate, disease, and habitat structure can all interact. These interactions can also produce a trophic cascade, where changes at one trophic level affect multiple levels below it. For example, adding or removing predators can change herbivore behavior, plant abundance, and even habitat structure. A common example involves wolves, elk, and plants. When large predators reduce herbivore pressure or change herbivore behavior, plants may recover in areas that were heavily browsed. This can affect birds, insects, stream banks, and other parts of the community. The exact details vary among ecosystems, but the general lesson is important: changing one species can indirectly affect many others.

Figure 12. Controls on Herbivores. Herbivore populations are shaped by both top-down forces, such as predators and disease, and bottom-up forces, such as plant abundance, plant quality, nutrients, and productivity. Changes at one trophic level can create indirect effects throughout the community.
Food Chains and Food Webs
A food chain is a simple pathway showing how energy and nutrients move from one organism to another. For example, grass is eaten by a grasshopper, the grasshopper is eaten by a frog, and the frog is eaten by a snake. Food chains are useful for teaching, but real communities are better represented as food webs. A food web is a network of feeding relationships. Most organisms eat more than one type of food, and most organisms are eaten by more than one type of consumer. Food webs include several trophic levels. Primary producers, such as plants, algae, and some bacteria, produce organic molecules using sunlight or chemical energy. Primary consumers eat producers. Secondary consumers eat primary consumers. Decomposers break down dead organisms and waste, returning nutrients to the environment. Food webs help ecologists understand indirect effects. If one predator declines, herbivores may increase. If herbivores increase, plants may decline. If plants decline, insects, birds, soil conditions, and stream habitats may also change. Community ecology is often about tracing these connections.
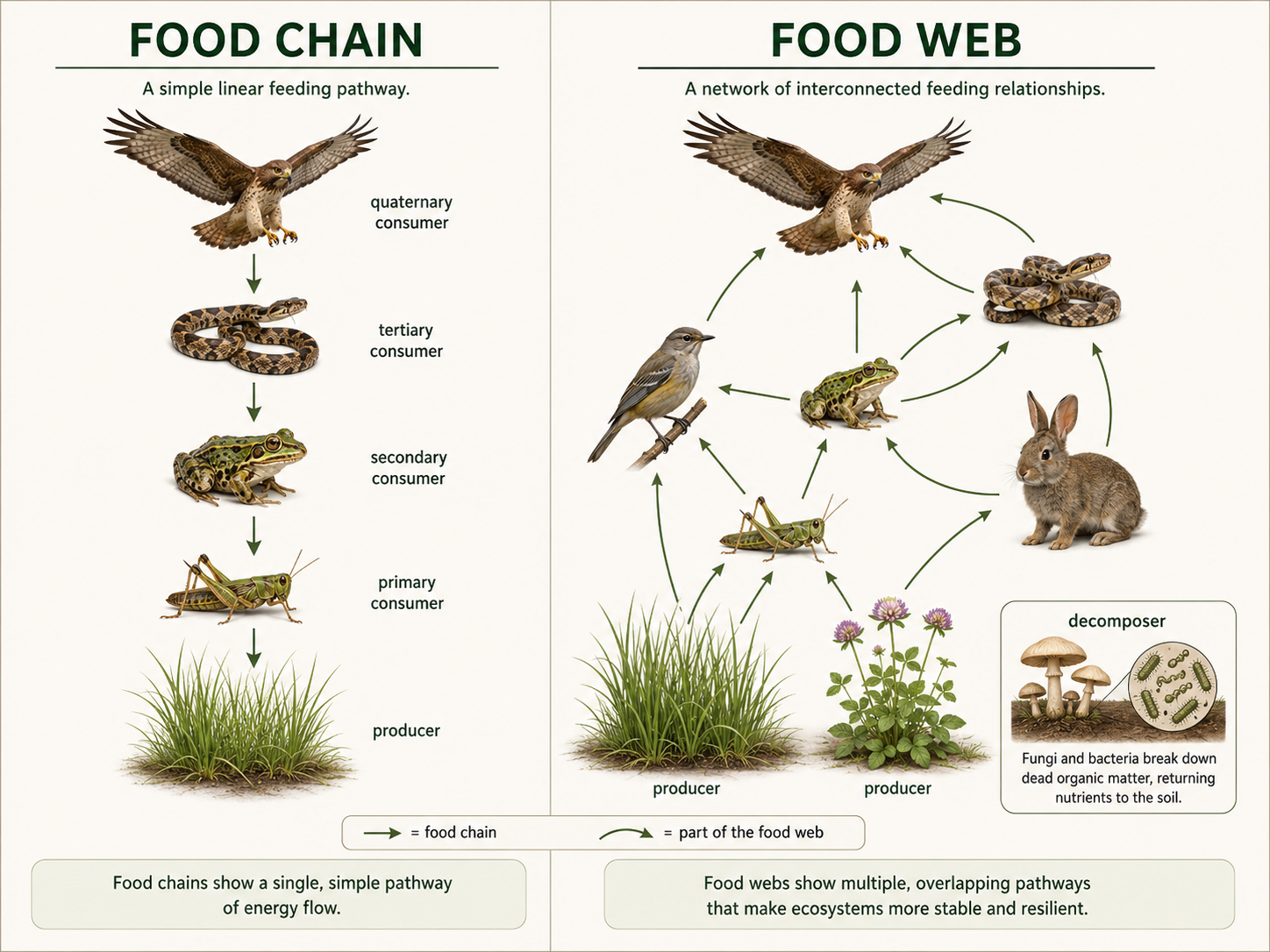
Figure 13. Food Chains and Food Webs. A food chain shows one pathway of energy and nutrient movement, while a food web shows many interconnected feeding relationships. Food webs better represent real communities because most organisms eat and are eaten by more than one
Keystone Species
A keystone species has an unusually large effect on its community relative to its abundance. Removing a keystone species can cause major changes in community structure. The term comes from the keystone in an arch. The keystone is not the largest stone, but it helps hold the arch together. Similarly, a keystone species may not be the most abundant species, but it plays a critical role in maintaining the community. Sea stars in rocky intertidal communities are a classic example. In some systems, predatory sea stars feed on mussels. Without the sea stars, mussels can dominate the space and exclude many other species. By limiting the mussels, sea stars help maintain diversity. Beavers can also function as keystone species or ecosystem engineers. By building dams, they create ponds and wetlands that provide habitat for fish, amphibians, insects, birds, and plants. Their behavior changes the physical environment in ways that affect many other species. Keystone species matter because they show that not all species have equal ecological influence. Losing one important species can reshape an entire community.

Figure 14. Keystone Species. A keystone species has a large effect on community structure relative to its abundance. In rocky intertidal communities, predatory sea stars can maintain diversity by preventing mussels from monopolizing space.
Ecosystem Engineers and Foundation Species
Ecosystem engineers are species that shape communities by changing the physical environment. Beavers are a clear example because their dams alter water flow and create wetlands. Earthworms, corals, oysters, prairie dogs, and burrowing animals can also engineer habitats. A foundation species is a species that strongly structures a community by creating habitat or providing major resources. Trees in a forest are foundation species because they create shade, leaf litter, nesting sites, food, and physical structure. Coral species build reefs that support thousands of other organisms. Kelp creates underwater forests that provide food and shelter for fish, invertebrates, and other marine life. Keystone species, ecosystem engineers, and foundation species are not always separate categories. A species can fit more than one category. What matters is that some species strongly influence the conditions under which many other species live.

Figure 15. Ecosystem Engineers and Foundation Species. Ecosystem engineers shape communities by changing the physical environment, while foundation species strongly structure communities by creating habitat or providing major resources. Some species may fit more than one ecological category.
Mutualism
Mutualism is an interaction in which both species benefit (+/+). Mutualisms are common and ecologically important, but they are not acts of kindness. Each species benefits because the interaction increases its survival, growth, or reproduction. One major example is the relationship between plants and mycorrhizal fungi. The fungi grow in or around plant roots and help plants absorb water and nutrients, especially phosphorus. In return, the plant provides the fungi with sugars produced by photosynthesis. Most land plants form some type of mycorrhizal association. Another example is pollination. Flowering plants often depend on animals such as bees, butterflies, birds, bats, or flies to move pollen between flowers. The plants gain reproduction, while pollinators gain nectar, pollen, or other rewards. Lichens are partnerships between fungi and photosynthetic partners, such as green algae or cyanobacteria. The photosynthetic partner produces sugars, while the fungus provides structure, protection, and access to water and minerals. Lichens are important pioneer organisms in some harsh environments because they can grow on rock surfaces and help begin soil formation. Some ants and acacia trees also form mutualisms. The tree provides food and shelter, while the ants defend the tree against herbivores and sometimes remove competing vegetation. Cleaner shrimp and cleaner fish provide another example. They remove parasites from larger fish. The cleaner gains food, and the client fish gains parasite removal. Mutualisms can be obligate, meaning one or both species depend on the interaction to survive or reproduce, or facultative, meaning the interaction is helpful but not strictly required. Some mutualisms are highly specialized, while others are flexible. Mutualisms can also break down. If one partner receives benefits but stops providing benefits in return, it may become a cheater.

Figure 16. Mutualism. Mutualism is a species interaction in which both partners benefit. Examples include plant-mycorrhizal fungus associations, pollination, lichens, ant-acacia interactions, and cleaner organisms that remove parasites from other animals.
Facilitation
Facilitation occurs when one species makes the environment more suitable for another species. Facilitation is especially important in stressful environments, such as deserts, salt marshes, rocky shores, and alpine habitats. For example, a shrub in a desert may provide shade that reduces soil temperature and water loss. Seedlings of other plants may survive better under the shrub than in open sun. In this case, the shrub facilitates the establishment of other species. In primary succession, early colonists may facilitate later species by building soil, adding organic matter, trapping moisture, or reducing harsh conditions. Lichens and mosses can help break down rock and contribute to early soil development. Grasses and small plants can stabilize soil and make it easier for shrubs or trees to establish later. Facilitation is important because not all interactions are competitive or consumer-based. Sometimes one species improves conditions for another, especially when the physical environment is difficult.

Figure 17. Facilitation. Facilitation occurs when one species makes the environment more suitable for another species. It is especially important in stressful environments and during succession, where early colonists may improve conditions for later species.
Disturbance
A disturbance is an event that changes a community by removing organisms, altering resource availability, or changing physical conditions. Disturbances can be small or large, frequent or rare, mild or severe. Examples include fires, hurricanes, floods, droughts, volcanic eruptions, landslides, disease outbreaks, storms, tree falls, and human activities such as farming, logging, development, and pollution. Disturbance reduces biomass, but it can also create new opportunities. For plants, disturbance may increase light, space, and nutrient availability. For animals, disturbance may remove shelter, reduce food, or create new habitat depending on the species involved. Disturbance is not necessarily “bad” for ecosystems. Many communities are adapted to, and even require, periodic disturbance. Some grasslands, pine forests, wetlands, and fire-adapted forests depend on disturbance to maintain their structure and diversity. A small disturbance, such as a tree falling in a forest, may create a gap where sunlight reaches the forest floor. This can allow seedlings, shrubs, and herbs to grow. A large disturbance, such as a severe hurricane, may reshape an entire forest. The ecological effects depend on the type of disturbance and the community’s history.

Figure 18. Disturbance. A disturbance is an event that changes a community by removing organisms, altering resources, or changing physical conditions. Disturbances can reduce biomass but also create openings for new growth and community change.
Disturbance Regimes
A disturbance regime is the typical pattern of disturbance in a particular ecosystem, including type, frequency, intensity, severity, size, and seasonality of disturbance. Fire is a good example. Some ecosystems experience frequent, low-intensity fires that burn grasses and leaf litter but leave many large trees alive. Other ecosystems experience rare, high-severity fires that kill most aboveground vegetation. Fire suppression has changed many North American ecosystems. The Smokey Bear campaign helped teach the public to prevent human-caused wildfires, but widespread fire suppression also interrupted natural fire regimes in many fire-adapted systems. In some forests and grasslands, decades without fire allowed fuel to build up, changed species composition, and reduced regeneration of fire-adapted plants. Many fire-adapted plants benefit from periodic fire because fire can reduce competition, release nutrients, open space, expose mineral soil, or trigger seed release and germination. Some pine species release seeds after fire. Many grassland plants regrow from underground tissues after burning. In these ecosystems, fire is not just a destructive event. It is part of the ecological rhythm. The important point is that each ecosystem has a disturbance history, and changing that history can change the community. Too much disturbance can prevent communities from recovering, but too little disturbance can also reduce diversity in systems adapted to regular disturbance.

Figure 19. Disturbance Regimes. A disturbance regime is the typical pattern of disturbance in an ecosystem. Fire frequency, intensity, severity, size, and seasonality can shape species composition, regeneration, and long-term community structure.
The Intermediate Disturbance Hypothesis
The intermediate disturbance hypothesis proposes that species diversity may be highest at intermediate levels of disturbance. When disturbance is very low, strong competitors may dominate and exclude other species. When disturbance is very high, only a few species can tolerate frequent disruption. At intermediate disturbance levels, both early and late successional species may coexist. For example, occasional tree falls in a forest can create light gaps that allow shade-intolerant plants to grow. If no gaps ever form, shade-tolerant competitors may dominate. If the forest is disturbed too often or too severely, many slow-growing species may not survive. This hypothesis is useful, but it is not a universal rule. Some ecosystems fit the pattern better than others. Modern ecologists treat it as one possible explanation for diversity, not as a law that applies everywhere. The effect of disturbance depends on the type of disturbance, the species present, the climate, the soil, and the history of the community.

Figure 20. Intermediate Disturbance Hypothesis. The intermediate disturbance hypothesis proposes that diversity may be highest at intermediate disturbance levels because both early and late successional species can coexist. The hypothesis is useful but does not apply equally to all ecosystems.
Succession
Ecological succession is the process of community change following disturbance or the creation of new habitat. Succession describes how species arrive, establish, interact, and replace one another over time. Primary succession begins in an environment where soil is absent or nearly absent. This can occur after a volcanic eruption, glacial retreat, landslide, or the formation of new rock surfaces. Early colonists must tolerate harsh conditions and may help build soil over time. Secondary succession occurs after a disturbance removes some organisms but leaves the soil mostly intact. Examples include abandoned farm fields, forests recovering after fire, areas recovering after hurricanes, and communities returning after moderate flooding. Secondary succession is usually faster than primary succession because soil, seeds, roots, spores, microbes, and surviving organisms may remain. Even when aboveground vegetation is removed, belowground roots, seed banks, and soil organisms can speed recovery. Succession is not simply plants replacing plants. Animals, fungi, bacteria, and other organisms also change during succession. As plant structure changes, habitat changes. Early successional habitats may support species that need open ground and sunlight. Later successional habitats may support species that need shade, woody debris, leaf litter, or complex vertical structure.

Figure 21. Succession. Ecological succession is the process of community change after disturbance or new habitat formation. Primary succession begins with little or no soil, while secondary succession occurs where soil and some biological materials remain.
Successional Communities
Early successional species are often called pioneer species, and are usually good dispersers, grow quickly, reproduce early, and tolerate harsh conditions. They may be small, short-lived, and poor competitors once larger or longer-lived species become established. Classic pioneer species include lichens, mosses, grasses, annual plants, and fast-growing shrubs or trees, depending on the ecosystem. In primary succession, lichens and mosses may be among the earliest colonists because they can tolerate exposed surfaces and low nutrient availability. In secondary succession, grasses, herbs, and fast-growing shrubs often appear early because soil is already present. Late successional species tend to grow more slowly, live longer, and compete effectively for resources such as light, water, and nutrients. They often invest more energy in survival, structure, defense, and long-term growth. Pioneer species are historically known as r-selected species and late successional species as K-selected species. These terms can still be useful as a simple teaching model, but modern ecologists recognize that life history strategies are more complex. Not all species fit neatly into these two categories. Succession can be influenced by three major processes: facilitation, inhibition, and tolerance. In facilitation, early species improve conditions for later species. In inhibition, early species prevent later species from establishing. In tolerance, later species are able to grow under conditions created by earlier species, but they are not necessarily helped by them. Succession is shaped by dispersal, soil development, climate, disturbance history, species interactions, chance events, and human activity.

Figure 22. Successional Communities. Successional communities change as species arrive, establish, interact, and replace one another over time. Early successional species often disperse and grow quickly, while late successional species tend to grow more slowly and compete effectively under established conditions.
Climax Communities and the Modern View of Succession
Frederick Clements, an influential early ecologist, proposed that communities develop through predictable stages toward a stable climax community. In this view, pioneer species are replaced by herbaceous plants, then shrubs and young trees, and eventually long-lived tree species. Once the climax community forms, it tends to maintain itself until another disturbance occurs. This model was important historically, but modern ecology treats it as too simple. Many communities do not move toward a single predictable endpoint. Instead, succession can follow multiple pathways depending on disturbance severity, climate, soil conditions, species arrivals, invasive species, and chance events. Communities are often mosaics of patches at different successional stages. A forest may contain old trees, young gaps, recovering areas, dead wood, and disturbed patches all at the same time. Disturbance is not an exception to community life; it is one of the forces that helps shape community structure. The climax community concept is still useful as a historical model, but students should understand that real ecosystems are dynamic, variable, and continually changing. A better modern question is not “What final community will this become?” but “What factors influence the pathway of change?”

Figure 22. Successional Communities. Successional communities change as species arrive, establish, interact, and replace one another over time. Early successional species often disperse and grow quickly, while late successional species tend to grow more slowly and compete effectively under established conditions.
Island Biogeography
The theory of island biogeography explains patterns of species richness on islands. It was developed by Robert MacArthur and E. O. Wilson and remains one of the most influential ideas in ecology. The theory proposes that the number of species on an island depends on a balance between immigration and extinction. Immigration is the arrival of new species. Islands closer to a mainland or other source of colonists usually receive more immigrants than isolated islands. Extinction is the loss of species from the island. Smaller islands usually have higher extinction rates because they have smaller populations, fewer resources, and less habitat diversity. Larger islands usually support more species because they contain more space, larger populations, and more habitat types. The theory predicts that large, nearby islands should have the most species, while small, isolated islands should have the fewest species. Island biogeography applies not only to oceanic islands but also to “habitat islands,” such as mountaintops, lakes, forest fragments, parks, and nature preserves surrounded by human development. This makes the theory important for conservation biology. Habitat fragmentation can isolate populations, reduce immigration, increase extinction risk, and lower biodiversity. For example, a small forest patch surrounded by roads, neighborhoods, and farms can function like an island. Species that cannot cross the surrounding landscape may become isolated in the patch. Over time, small isolated populations are more likely to disappear. Conservation biologists use this idea when designing wildlife corridors, protected areas, and habitat restoration projects.
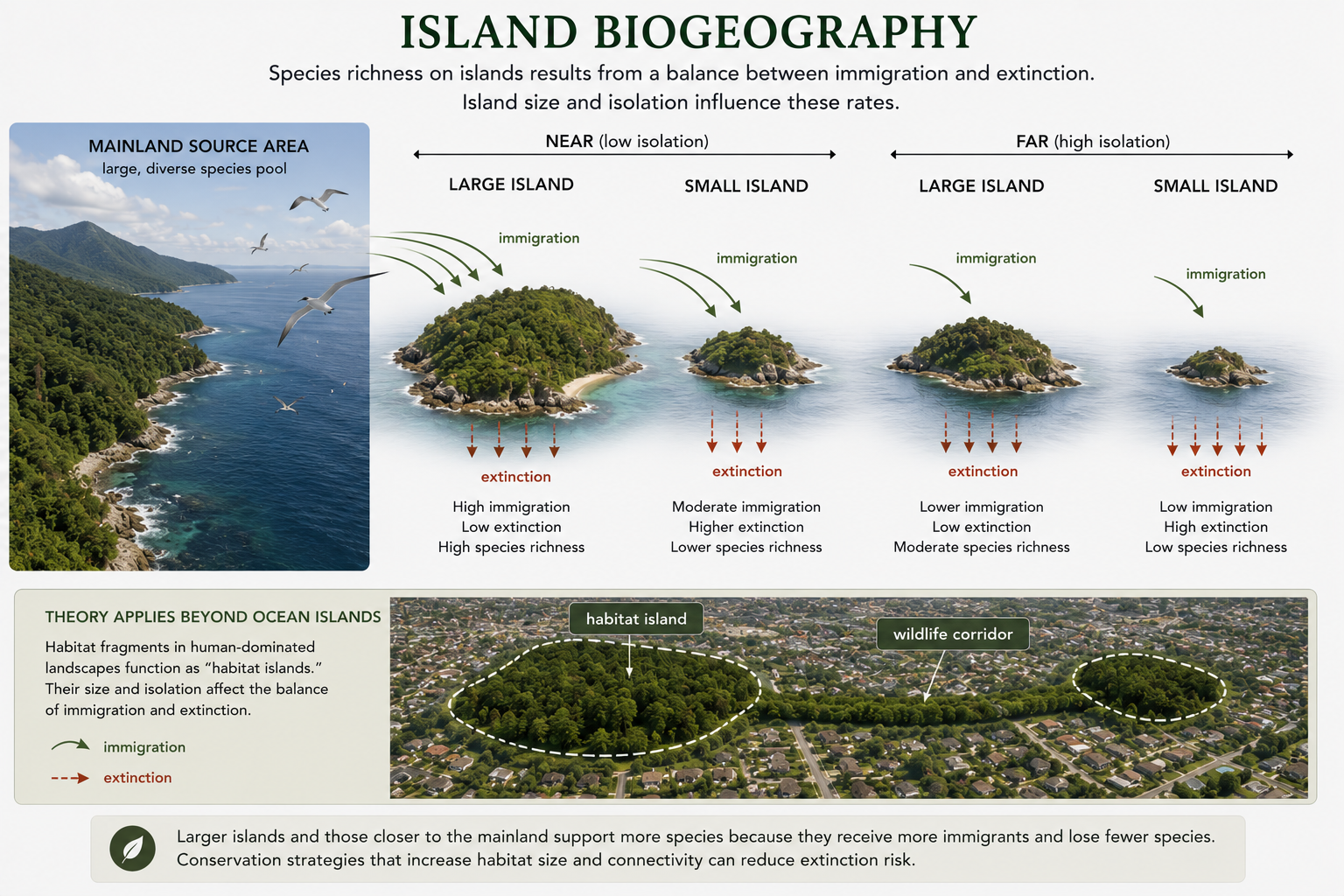
Figure 24. Island Biogeography. Island biogeography explains species richness as a balance between immigration and extinction. Large, nearby islands are predicted to support more species, while small, isolated islands usually support fewer species.
Community Ecology and Biodiversity
Community ecology helps explain why biodiversity is distributed unevenly across Earth. Some communities, such as tropical rainforests and coral reefs, contain extremely high species richness. Others, such as deserts, tundra, or highly disturbed habitats, may contain fewer species. Biodiversity is influenced by climate, productivity, habitat complexity, disturbance, species interactions, evolutionary history, and geographic isolation. No single factor explains all patterns. Ecologists often distinguish between different kinds of diversity. Species richness is the number of species in a community. Species evenness describes how evenly individuals are distributed among species. A community with ten species may be dominated by one species, or it may have similar numbers of individuals in each species. Species diversity combines richness and evenness. These measures matter because communities with higher diversity may be more resilient, provide more ecosystem services, and contain more complex networks of interactions. However, diversity alone does not guarantee stability. The identities of species and the roles they play also matter.

Figure 25. Community Ecology and Biodiversity. Biodiversity varies among communities and is influenced by climate, productivity, habitat complexity, disturbance, species interactions, evolutionary history, and geographic isolation. Species richness counts the number of species, while evenness describes how evenly individuals are distributed among them.