
Chapter: Animals
Characteristics of Animals
All animals share a set of fundamental biological characteristics that distinguish them from other forms of life. Animals are multicellular, eukaryotic organisms whose cells lack cell walls, allowing for greater flexibility and specialization. They are heterotrophs, meaning they obtain energy and nutrients by consuming other organisms or organic matter rather than producing their own food. Animal cells are organized into specialized tissues that perform specific functions, and most animals possess muscle and nervous tissues that enable movement and responses to environmental stimuli. Animals typically reproduce sexually, with a diploid-dominant life cycle in which a fertilized egg develops through a series of embryonic stages, including a blastula. During development, cells differentiate into specialized structures and organ systems. Although some animals, such as sponges, lack true tissues and most adult animals are capable of movement, all animals share a common evolutionary origin and possess the genetic and developmental traits that unite them within the animal kingdom.
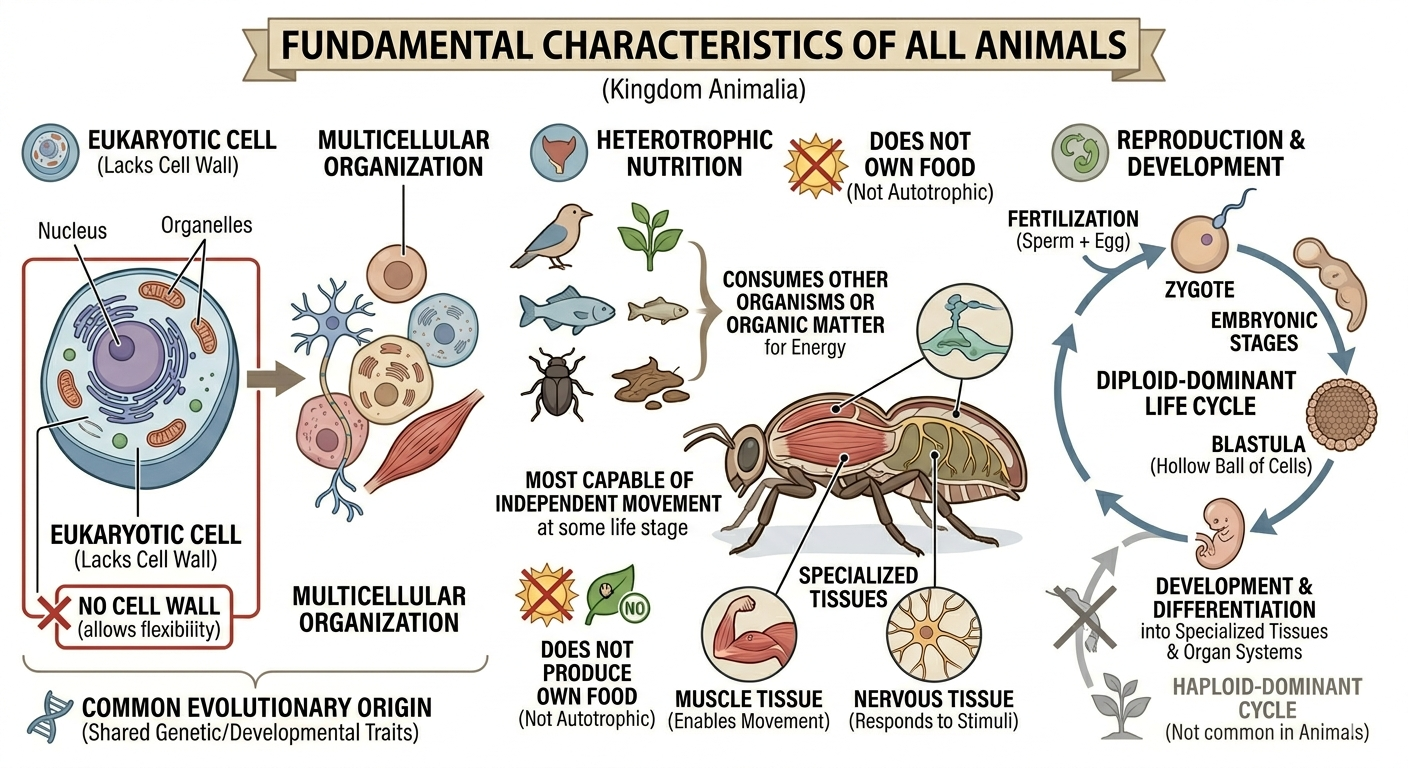
Figure 1. Characteristics of Animals. Animals are multicellular, heterotrophic eukaryotes that lack cell walls and possess specialized tissues, including muscle and nervous systems. They share sexual reproduction with embryonic development through a blastula stage and originate from a common evolutionary ancestor, despite their wide diversity of forms and lifestyles.
Choanoflagellates: Proto-Animals
Choanoflagellates are single-celled protists that are widely regarded as the closest living relatives of animals. These microscopic organisms inhabit both marine and freshwater environments, where they feed by generating water currents with a whip-like flagellum. Surrounding the flagellum is a collar of microvilli that traps bacteria and other small food particles. Although choanoflagellates are unicellular, some species can form temporary colonies, providing clues about how multicellular life may have evolved. Genetic evidence indicates that animals and choanoflagellates share a recent common ancestor, and many of the genes involved in cell adhesion, communication, and signaling in animals are also found in choanoflagellates. These findings suggest that the molecular foundations of multicellularity evolved before the first animals appeared.
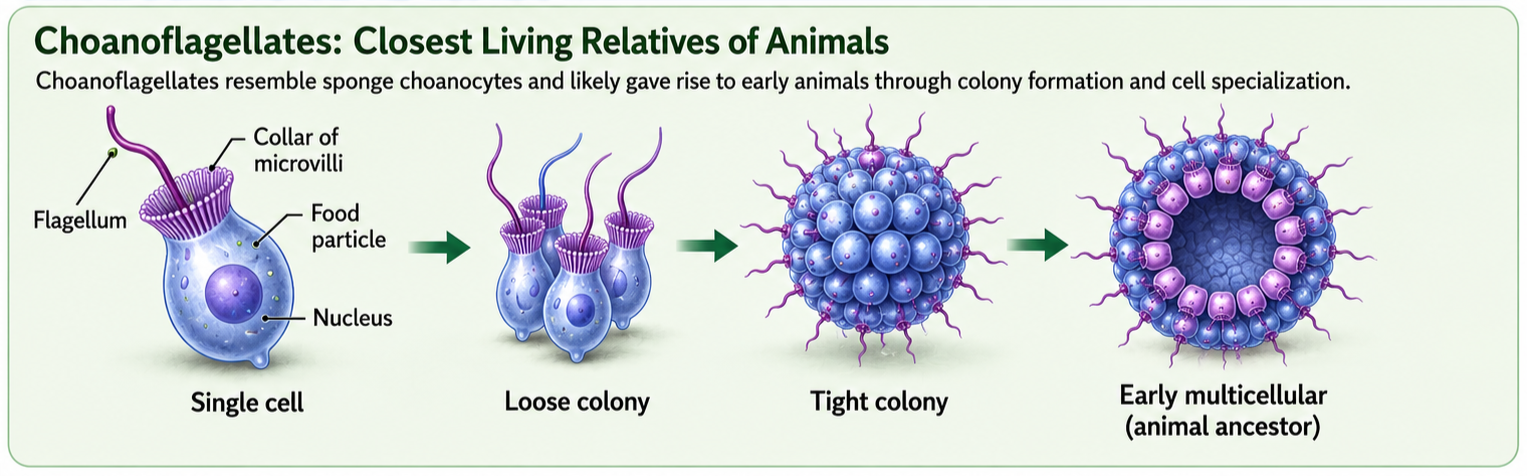
Figure 2. Choanoflaggelates. Choanoflagellates are single-celled protists and the closest living relatives of animals. Their feeding structure, colonial forms, and animal-like genes for cell adhesion and signaling provide clues about how multicellularity evolved before the first animals appeared.
Origin of True Animals
One leading hypothesis for the origin of animals proposes that the earliest animals evolved from a choanoflagellate-like ancestor. Rather than living as independent cells, these ancestral organisms may have begun forming stable colonies in which cells remained attached after division. Over time, natural selection favored greater cooperation among cells, leading to specialization and division of labor. Some cells became more efficient at feeding, while others provided structural support, reproduction, or coordination. This transition culminated in the evolution of simple multicellular organisms resembling modern sponges. Evidence for this relationship can still be seen today in sponge choanocytes, specialized feeding cells that closely resemble choanoflagellates in both structure and function. Like choanoflagellates, choanocytes use a flagellum to generate water currents and a collar of microvilli to capture food particles.
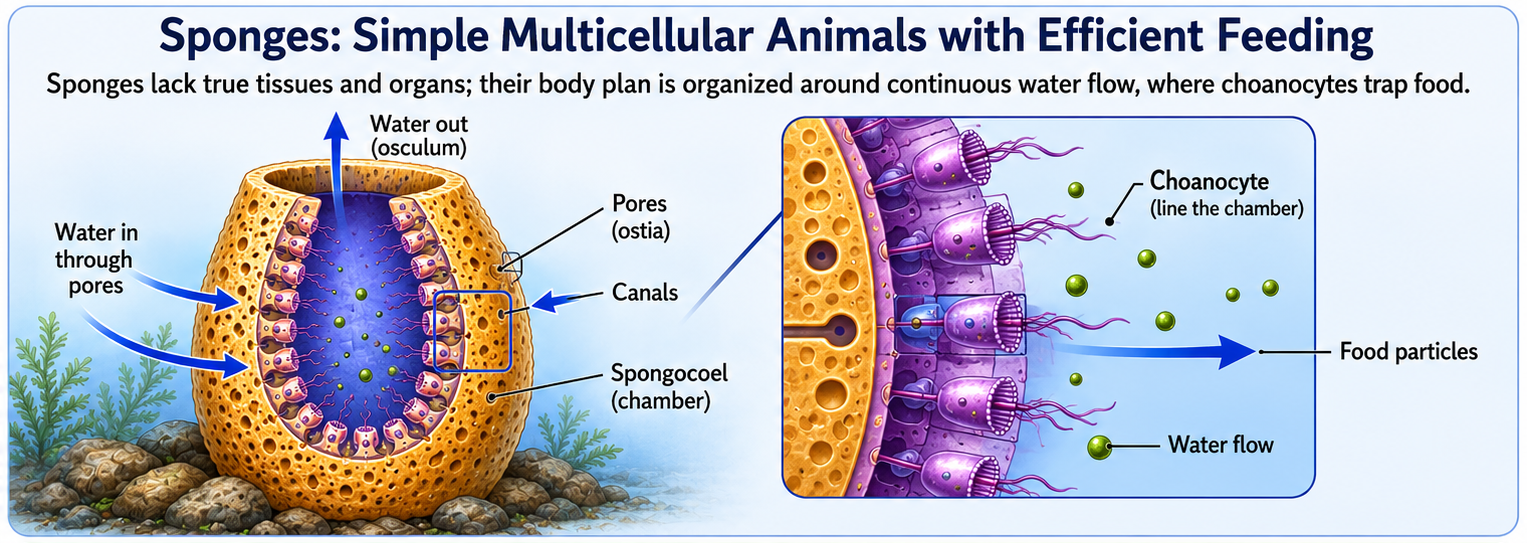
Figure 3. The Origin of Animals. One leading hypothesis proposes that animals evolved from choanoflagellate-like ancestors that formed stable colonies, eventually leading to cell cooperation, specialization, and simple multicellular bodies like sponges. Sponge choanocytes still show this connection because they use a flagellum and collar of microvilli to move water and capture food, much like choanoflagellates.
Phylum Porifera (Sponges)
Kingdom Animalia: Phylum Porifera
Sponges (Phylum Porifera) are among the simplest living animals and likely resemble some of the earliest animal forms. They lack true tissues, organs, muscles, and nervous systems, yet they are fully multicellular organisms composed of specialized cell types. Most sponges are sessile, remaining attached to rocks or other substrates throughout their adult lives. Their bodies are organized around a system of pores, canals, and chambers that continuously circulate water. As water flows through the sponge, choanocytes lining the internal chambers capture and digest suspended food particles. This arrangement allows thousands of feeding cells to work together, greatly increasing feeding efficiency compared to a single-celled ancestor. The sponge body plan represents one of the earliest examples of a multicellular animal design—a tube-within-a-tube organization that maximizes water flow, feeding efficiency, and the exchange of gases and wastes. Sponges also possess remarkable regenerative abilities and can often rebuild damaged tissues. Despite their simplicity, they represent a major evolutionary milestone because they demonstrate how specialized cells can cooperate within a multicellular organism, laying the foundation for the evolution of more complex animal body plans.
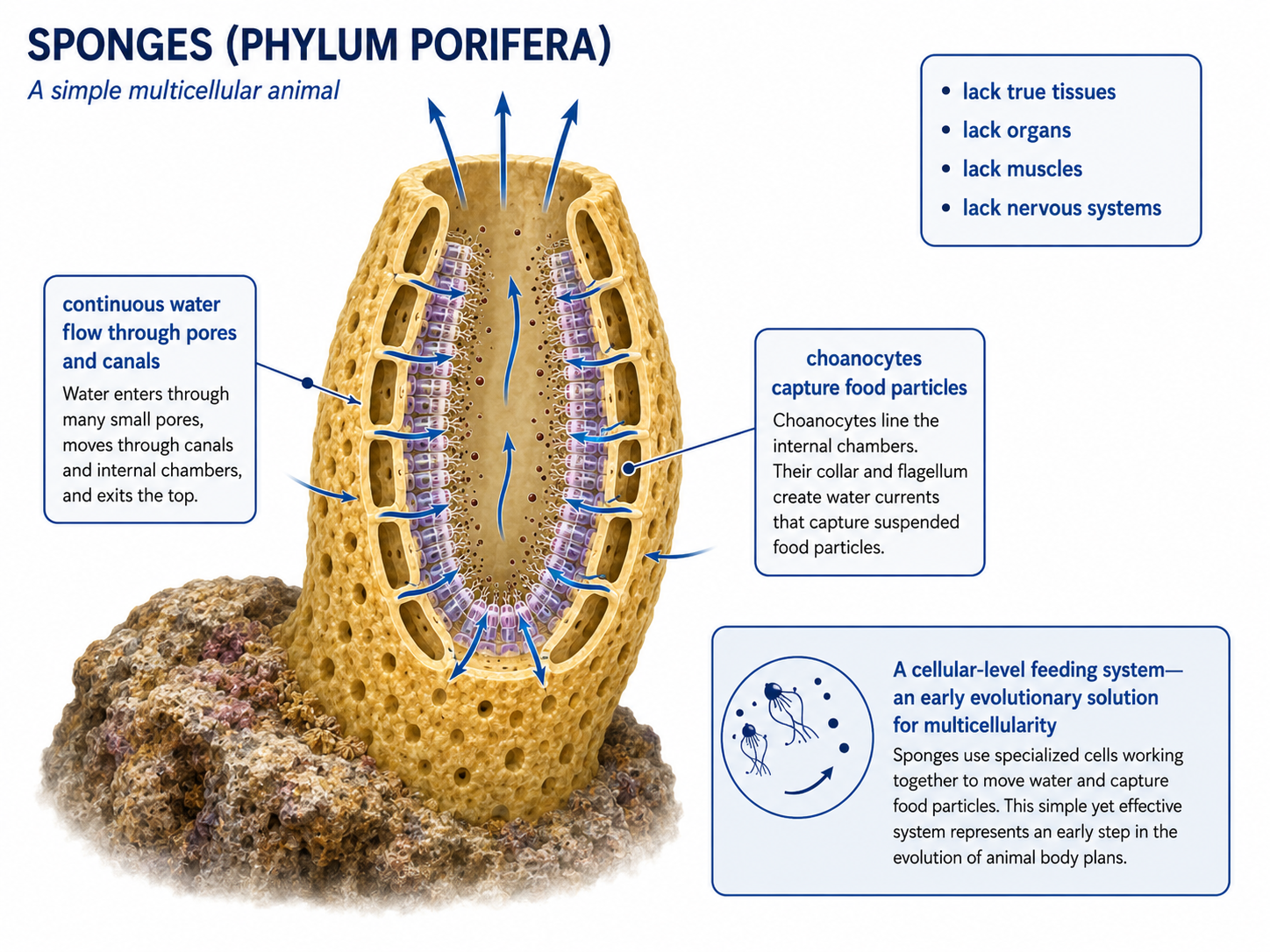
Figure 4. Phylum Porifiera. Sponges are simple multicellular animals that lack true tissues, organs, muscles, and nervous systems. Their body plan is organized around continuous water flow through pores and canals, where choanocytes capture food particles. This efficient cellular-level feeding system represents an early evolutionary solution for multicellularity and provides insight into the origin of animal body plans.
Eumetazoa: The Origin of True Tissues
Kingdom Animalia: Eumetazoa
Eumetazoa is the clade of animals with true tissues. This group includes nearly all animals: ctenophores, cnidarians, and bilaterians. Eumetazoa excludes sponges, which have specialized cells but lack true tissues and organs. Sponges are multicellular animals, but their cells are only loosely organized and do not form true tissues. The next major step in animal evolution was the origin of true tissues: organized groups of specialized cells that are physically connected, communicate with one another, and function together as coordinated units. This innovation allowed cells to become more specialized and greatly increased the complexity and efficiency of animal body plans. Below are some of the key emergent structures that allowed animals to continue to divers The evolution of true tissues allowed the development of structures such as epithelial tissue, which covers body surfaces and lines internal cavities. The cells within epithelial tissues are tightly connected by protein structures called tight junctions, forming protective barriers that regulate the movement of substances and help maintain internal conditions. The appearance of tissues enabled cells to work together as integrated systems for protection, feeding, digestion, and basic nervous system function, representing a major advance over the simpler organization seen in sponges.
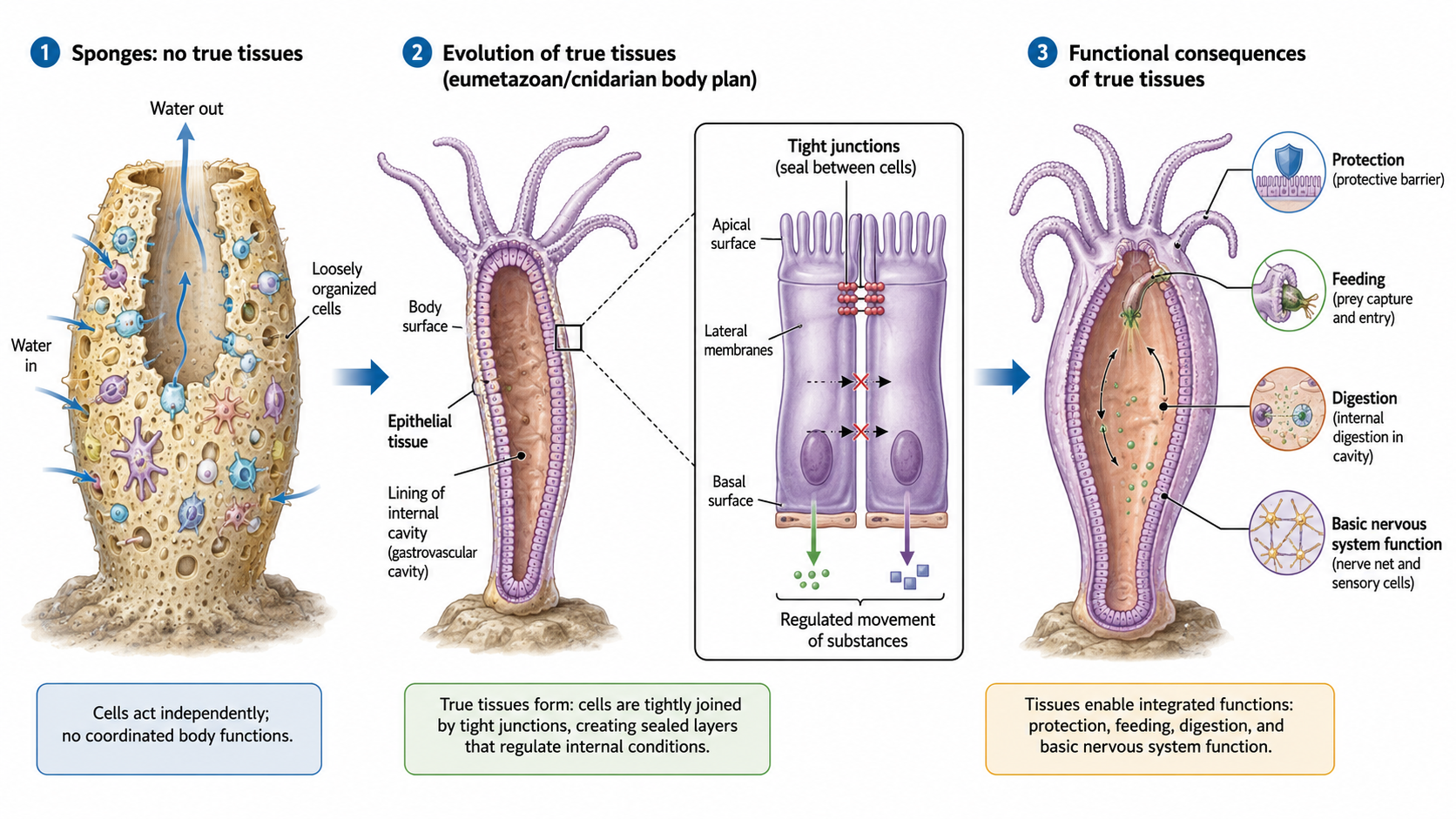
Figure 6. Evolution of true tissues in animals. Sponges lack true tissues and have loosely organized cells. In early animals with true tissues, epithelial cells became tightly joined by tight junctions, forming protective layers that regulate movement of substances. These tissues allowed cells to work together for protection, feeding, digestion, and basic nervous system function.
Diploblasts: The Origin of Germ Layers
Kingdom Animalia: Eumetazoa: Radiata
A major division in animal evolution is based on embryonic development, specifically the number of germ layers that form during early development. Germ layers are groups of embryonic cells that give rise to all of the tissues and organs of the adult animal. Diploblasts are the simplest animals with true tissues and develop from two germ layers: the ectoderm ("outer layer") and the endoderm ("inner layer"). Diploblasts are in the clade: Radiata. The ectoderm forms the outer body covering and gives rise to structures involved in interacting with the environment, including the skin and nervous system. The endoderm forms the lining of the digestive cavity, where food is digested and nutrients are absorbed. Because diploblasts possess only two germ layers, their body organization is relatively simple compared to that of triploblastic animals, which possess an additional third germ layer called the mesoderm.
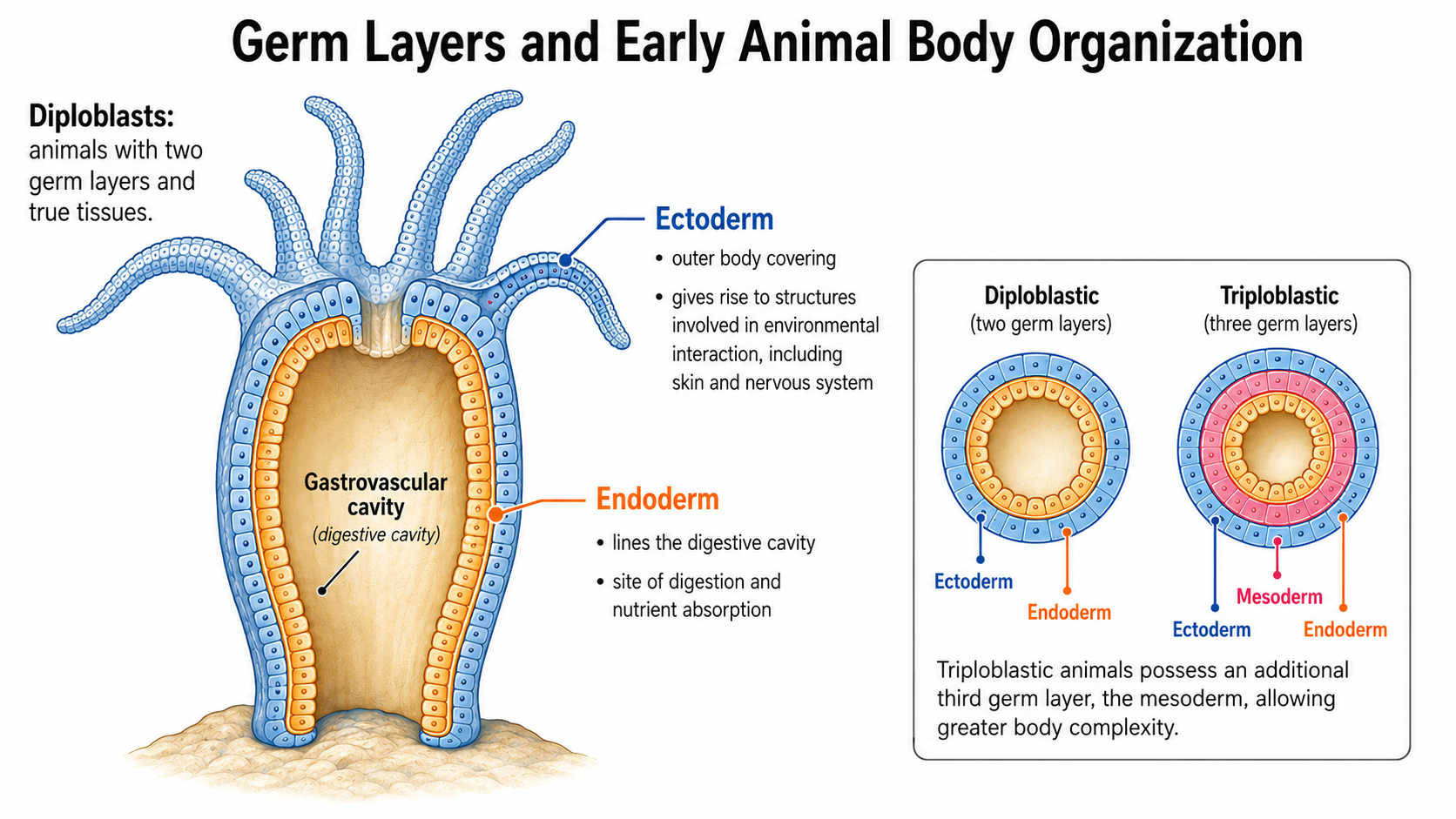
Figure 5. Germ layers in animal development. Diploblasts have two germ layers: an outer ectoderm and an inner endoderm. Triploblasts have a third layer, the mesoderm, which allows more complex body structures to develop.
Diploblasts: The Origin of Symmetry
Kingdom Animalia: Eumetazoa: Radiata
The transition from sponges to diploblasts was also accompanied by the evolution of more organized body symmetry. Most sponges are asymmetrical, meaning they lack a consistent body plan or plane of symmetry. Diploblasts, however, are generally radially symmetrical, with body parts arranged around a central axis and multiple planes of symmetry. This body plan is well suited for aquatic organisms that interact with their environment from all directions. Animals such as jellyfish, sea anemones, corals, and comb jellies can capture food and respond to stimuli regardless of the direction from which they arrive, making radial symmetry advantageous for drifting, floating, or sessile lifestyles.
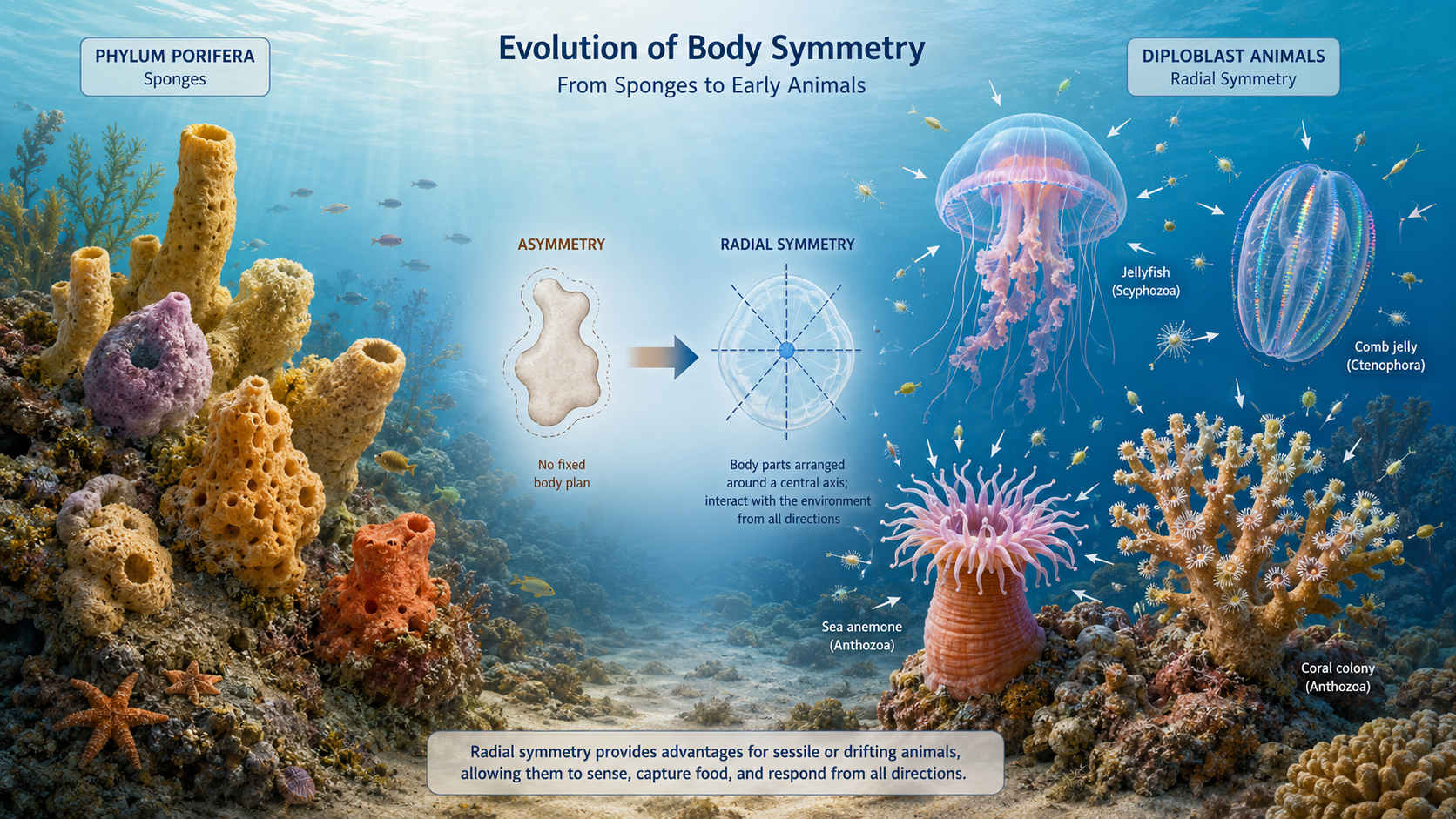
Figure 7. Origin of Symmetry. Evolution of body symmetry from asymmetrical sponges to radially symmetrical diploblast animals. Sponges lack a fixed body plan, while diploblasts such as jellyfish, sea anemones, corals, and comb jellies have body parts arranged around a central axis, allowing them to sense, capture food, and respond to the environment from multiple directions.
Diploblast Diversity
Kingdom Animalia: Eumetazoa: Radiata
Diploblasts include two major phyla:
Phylum Cnidaria (jellyfish, corals, and sea anemones)
Phylum Ctenophora (comb jellies).
Together, these groups represent an important evolutionary transition between the simple cellular organization of sponges and the more complex body plans of triploblastic animals.
Phylum Cnidaria (Jellyfish, Corals and Anemones)
Kingdom Animalia: Eumetazoa: Radiata: Phylum Cnidaria
Cnidarians (Phlylum Cnidaria) include jellyfish, corals, and sea anemones and are defined by their relatively simple body organization and specialized stinging cells, known as cnidocytes. Most cnidarians exhibit radial symmetry, meaning their body parts are arranged around a central axis, which suits a lifestyle where they interact with the environment from all directions. A key feature of the group is the presence of two distinct life stages in many species: a free-swimming medusa and a sessile polyp. The medusa stage, best represented by jellyfish, is typically bell-shaped and moves through the water by rhythmic contractions of its body, allowing it to drift or actively swim through open ocean environments. These medusae possess tentacles lined with cnidocytes, specialized cells that contain harpoon-like structures capable of injecting toxins, which are used to immobilize prey and for defense. In contrast, the polyp stage is usually anchored to a surface such as rock, coral reef, or other hard substrate. Polyps are often cylindrical and oriented with their mouth and tentacles facing upward, allowing them to capture passing food particles or small organisms. Many polyps reproduce asexually through budding, where new individuals grow directly from the parent organism and eventually detach or remain connected to form colonies. In colonial cnidarians such as corals, this process contributes to the formation of large reef structures that provide habitat for diverse marine life.
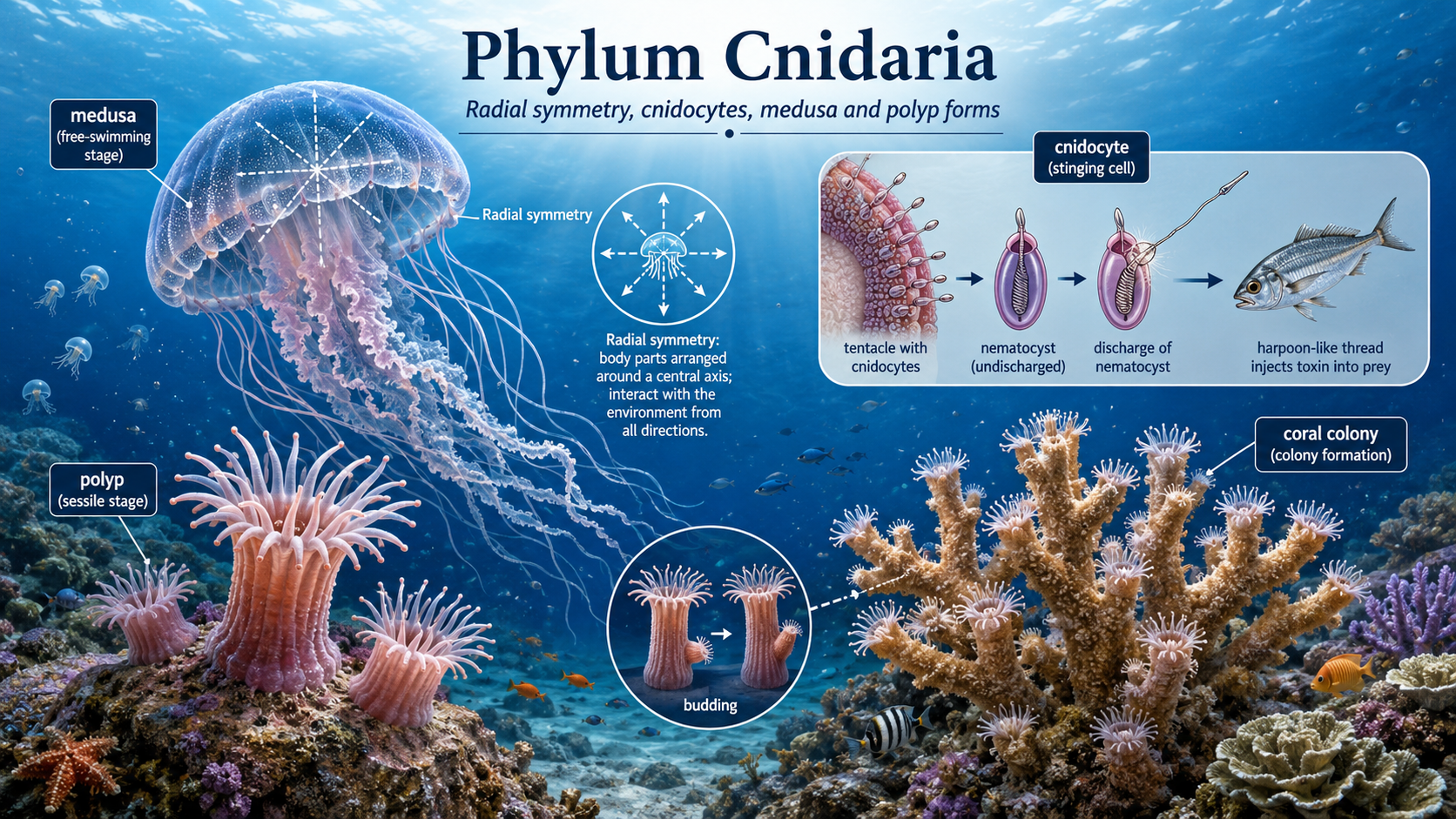
Figure 8. Phylum Cnidaria. Cnidarians include jellyfish, sea anemones, and corals, all of which share radial symmetry and specialized stinging cells called cnidocytes. Many species alternate between a free-swimming medusa stage and a sessile polyp stage, while colonial forms such as corals grow by budding and can build reef habitats that support diverse marine life.
Phylum Ctenophora (Comb Jellies)
Kingdom Animalia: Eumetazoa: Radiata: Phylum Ctenophora
Ctenophores, commonly known as comb jellies, resemble jellyfish in overall appearance but belong to a distinct evolutionary lineage separate from cnidarians. They are typically transparent and gelatinous, with a delicate body plan adapted for drifting and slow swimming in marine environments. Instead of stinging cells, ctenophores capture prey using sticky cells called colloblasts, which adhere to plankton and small organisms. Their movement is driven by eight rows of fused cilia arranged in comb-like bands along the body surface. These cilia beat in coordinated waves, producing a shimmering, rainbow-like effect as light diffracts off their movement. This ciliary locomotion allows ctenophores to glide smoothly through the water while feeding, often in a slow but efficient drift through plankton-rich waters. Pro-tip: Bioluminescent members of the Ctenophora can sometimes be observed glowing in the waters near Titusville, FL where gentle disturbances in the water trigger flashes of blue-green light as they drift through plankton-rich coastal habitats. The best time to go is summer months on a new moon and clear skies.

Figure 9. Phylum Ctenophora. Ctenophores, or comb jellies, are transparent gelatinous marine animals in Phylum Ctenophora that move using eight rows of fused cilia called comb plates. Unlike cnidarians, they lack stinging cells and instead capture plankton with sticky colloblasts, while some species can produce blue-green bioluminescent flashes when disturbed.
Bilateria: Bilateral Symmetry, Triploblasty, Cephalization
Kingdom Animalia: Eumetazoa: Bilateria
All other major animal groups are bilaterally symmetrical (Clade Bilateria). In additioni, members of Bilateria are triploblasts, meaning they possess a third germ layer called the mesoderm. This additional layer allows for the development of more complex organs and body systems, supporting the wide diversity of structures and functions seen across the animal kingdom today.
Bilateria: Triplobasty (Origin of the Mesoderm)
Kingdom Animalia: Eumetazoa: Bilateria
Triploblasts are animals that develop from three primary embryonic tissue layers: the ectoderm, mesoderm, and endoderm. The ectoderm forms the outer body covering (“the skin”) and contributes to the nervous system and sensory structures, while the endoderm gives rise to the lining of the digestive tract and associated internal organs, from mouth to anus. The key innovation in triploblasts is the mesoderm, the middle layer, which produces most internal body organs such as muscles, circulatory tissues, and many internal organs. This added layer allows for much greater body complexity, including the development of true organs and more efficient internal transport systems compared to simpler diploblastic animals.
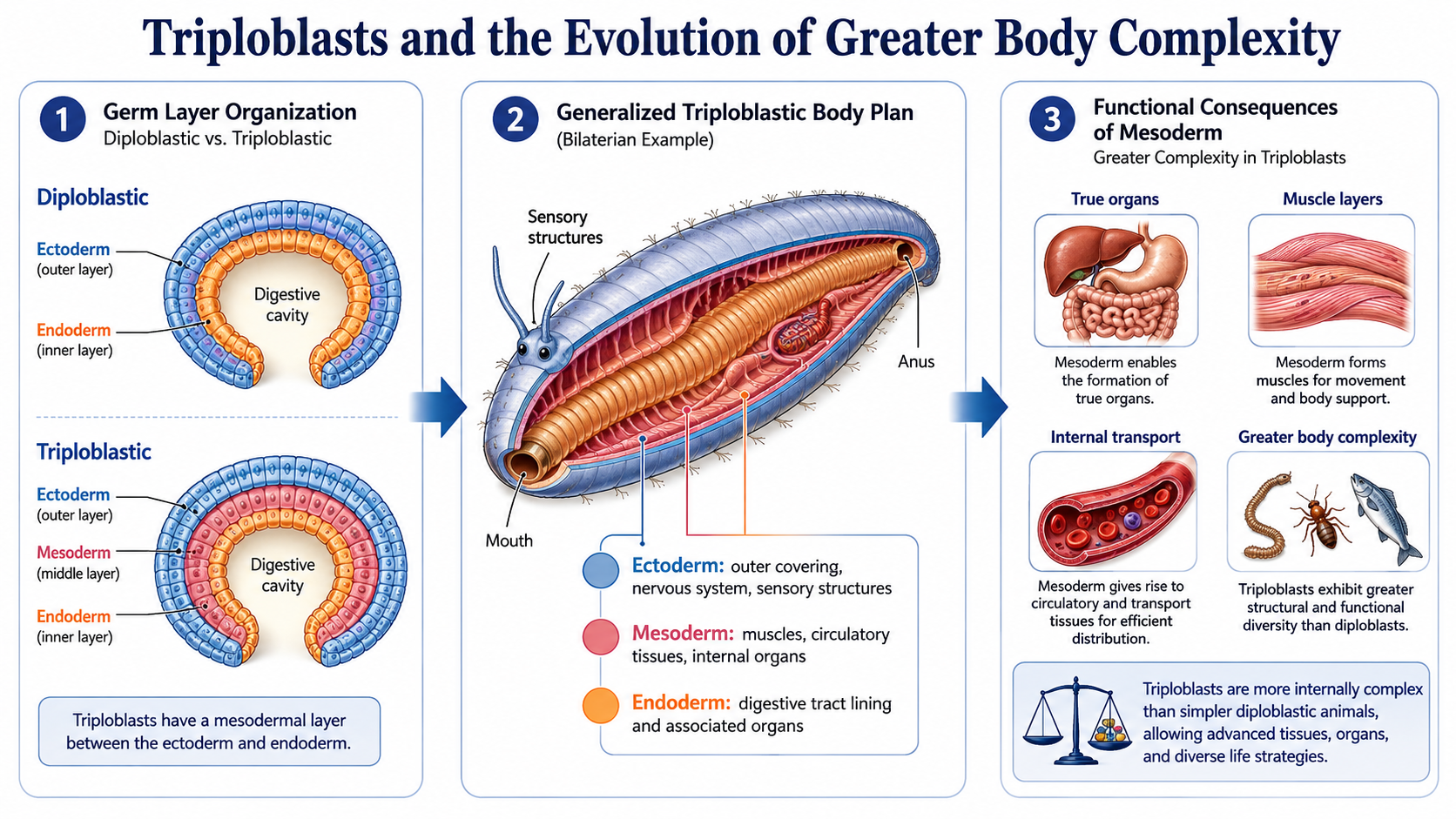
Figure 10. Triploblasts and greater body complexity. Triploblastic animals develop from three embryonic germ layers: ectoderm, mesoderm, and endoderm. The addition of the mesoderm allows the formation of muscles, circulatory tissues, internal organs, and more complex body systems, making triploblasts structurally and functionally more advanced than diploblastic animals.
Bilateria: Origin of Bilateral Symmetry and the Head Region
Kingdom Animalia: Eumetazoa: Bilateria
Triploblasts are also strongly associated with bilateral symmetry, meaning the body can be divided into mirror-image left and right halves along a single plane. This body plan is common in animals that move actively through their environment. Bilateral symmetry supports directional movement and leads to functional specialization of the anterior (front) end of the body. This forward-facing orientation is closely tied to cephalization, or the development of a head region, allowing a concentration of sensory structures, feeding appendages, and neural processing at the front end of the body. Cephalization provides a major evolutionary advantage because it allows organisms to process information and respond to stimuli more efficiently in the direction of movement. As a result, many bilateral animals develop a defined head region.]
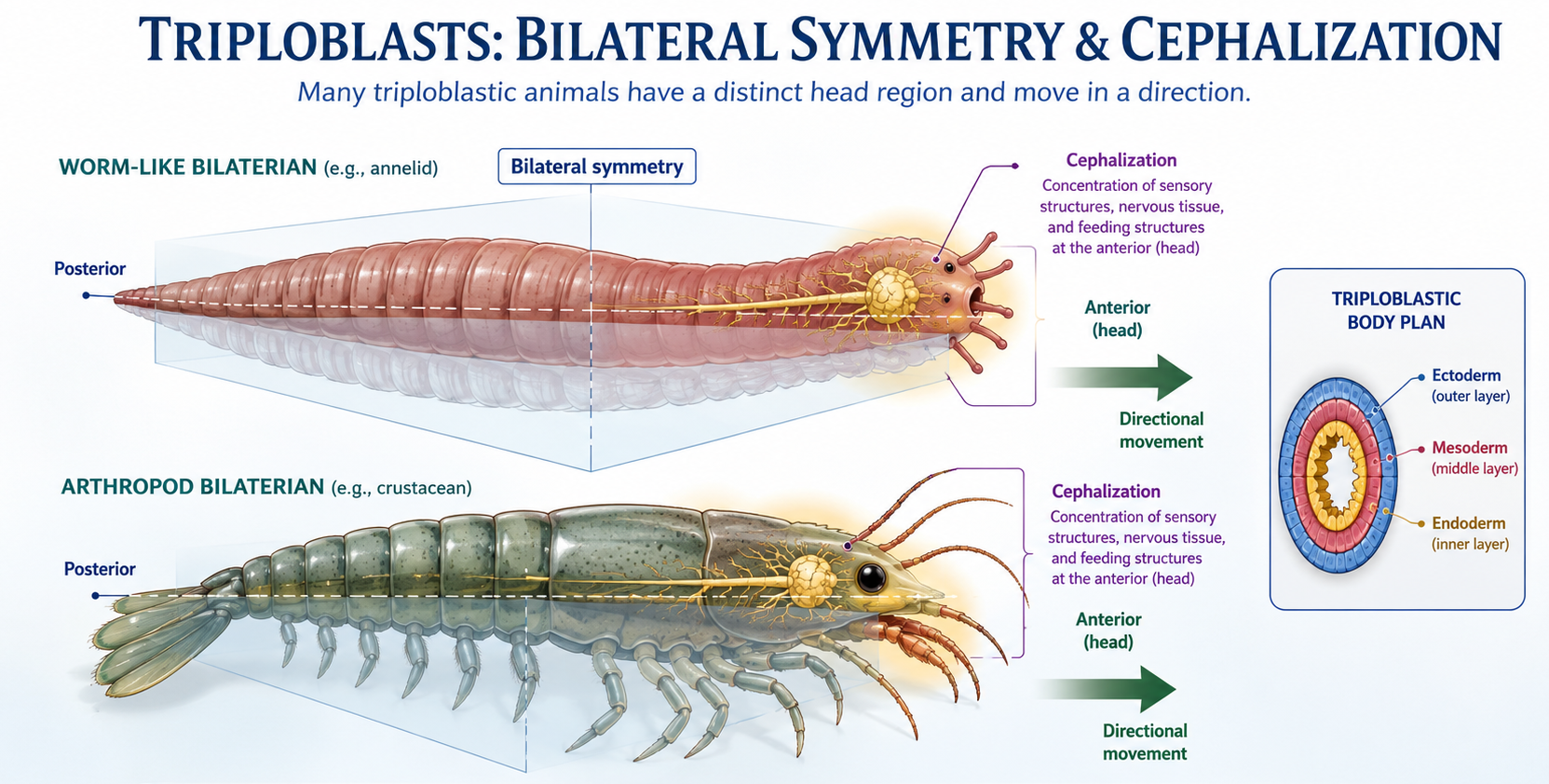
Figure 11. Bilateral symmetry and cephalization in triploblasts. Many triploblastic animals have left and right mirror-image body halves and move in a consistent forward direction. This body plan favors cephalization, with sensory structures, feeding structures, and nervous tissue concentrated at the anterior end to form a distinct head region.
Phylum Acoelomorpha
Kingdom Animalia: Eumetazoa: Bilateria: Phylum Acoelomorpha
Acoelomorphs (Phylum Acoelomorpha) are very simple bilaterian animals that help illustrate the transition from early diploblastic body plans to more complex triploblastic and coelomate body structures. Diploblastic animals, such as cnidarians, have two tissue layers and simple organization without true organs or a body cavity. Acoelomorphs have bilateral symmetry and a more advanced body plan, but they are still highly simplified compared to most bilaterians. They lack a true coelom (fluid-filled body cavity) and often lack a gut cavity, instead having a solid or partially digested internal region. This reduced internal compartmentalization makes them useful for understanding early steps away from diploblast-like organization. Acoelomorphs are not direct ancestors of coelomates such as annelids or vertebrates, but they represent an early-diverging bilaterian lineage that helps explain how complexity evolved. Their body plan shows early triploblastic features, including mesoderm tissue, but they do not form a true coelom, which in later animals supports organ organization, movement efficiency, and specialization. Overall, acoelomorphs represent an intermediate level of organization between diploblastic animals and more complex coelomates, showing how bilateral symmetry and early tissue layering likely evolved before fully developed body cavities appeared.
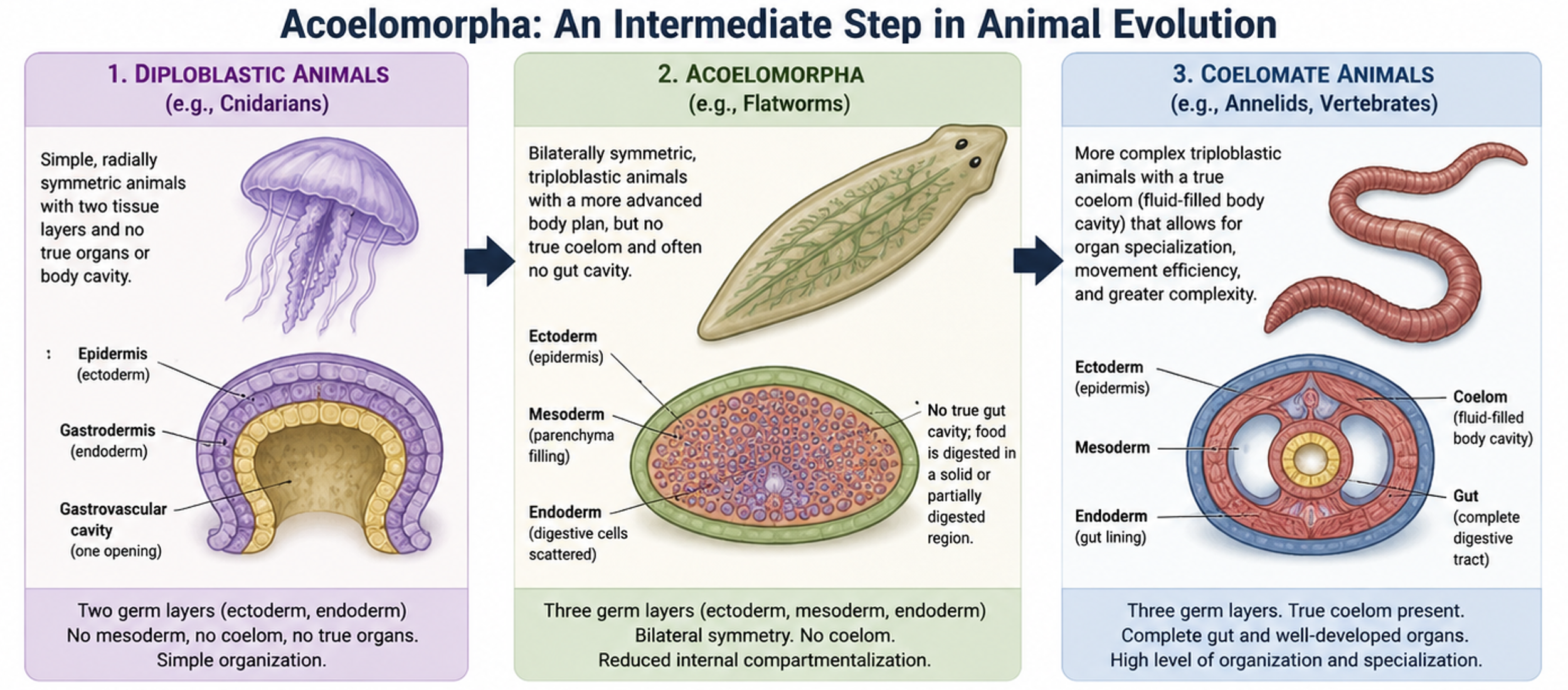
Figure 11. Acoelomorpha. Acoelomorphs are not direct ancestors of coelomates like annelids or vertebrates, but they are an early-diverging bilaterian lineage that helps explain how animal complexity evolved. They show early triploblastic traits, including mesoderm-like tissue, but lack a true coelom, which in later animals supports organ organization, movement efficiency, and specialization. Overall, they represent an intermediate level of organization between diploblastic animals and more complex coelomates.
Nephrozoa: Evolution of the Coelom
Kingdom Animalia: Eumetazoa: Bilateria: Nephrozoa
Early triploblastic animals such as Acoelomorpha are acoelomates, meaning they lack a coelom entirely. In these organisms, the space between the digestive tract and the outer body wall is filled with solid tissue rather than a fluid-filled cavity. In most other triploblastic animals, a key evolutionary innovation is the development of the coelom, a fluid-filled body cavity that forms entirely within the mesoderm and is completely lined by mesodermal tissue. This cavity allows internal organs to develop and move independently of the body wall and improves internal transport, support, and overall structural organization in more complex animals. These organisms are called coleomates, making up the clade Nephrozoa. Nephrozoa is split into to more clades: Protostomia and Deuterostomia.
Nephrozoa: Hydrostatic Movement
Kingdom Animalia: Eumetazoa: Bilateria: Nephrozoa
The origin of the coelom also plays an important role in movement in early coelomates, animals with a coelom. In these animals, like earthworms, the coelomic fluid acts as a hydrostatic skeleton, meaning it provides internal pressure that supports the body and allows movement. The body wall contains circular and longitudinal muscles surrounding the fluid-filled cavity. When circular muscles contract, the body becomes longer and thinner; when longitudinal muscles contract, the body becomes shorter and thicker. Because the fluid is incompressible, these coordinated muscle contractions change body shape rather than volume, producing a controlled wave-like motion. This allows worms to move efficiently through soil or water using peristaltic (wave-like) contractions along the length of the body.
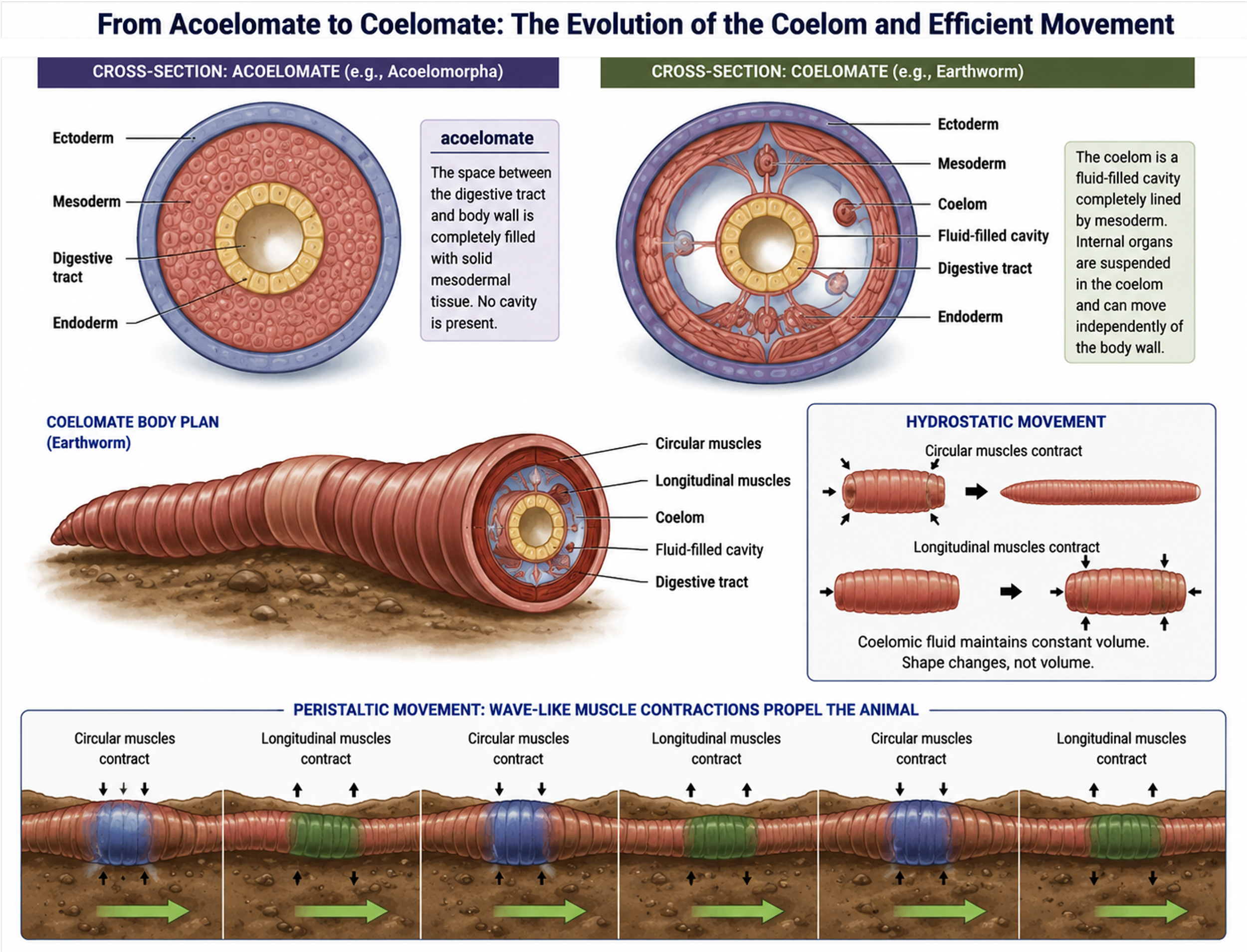
Figure 8. The Coelom. Early triploblastic animals lack a coelom and have solid mesodermal tissue between the body wall and gut. In coelomates, a fluid-filled cavity lined by mesoderm allows organs to move independently and enables hydrostatic movement. Coordinated muscle contractions acting on coelomic fluid produce efficient wave-like locomotion in animals such as earthworms.
Protostomes and Deuterostomes
Kingdom Animalia: Eumetazoa: Bilateria: Nephrozoa: Protostomia vs. Deuterostomia
Beyond sponges, cnidarians, ctenophores, and Acoelomorpha, most animals are coelomates are in the clade Nephrozoa and fall into one of two major evolutionary lineages: protostomes (Clade Protostomia) or deuterostomes (Clade Deuterostomia). These two groups make up the bulk of bilaterally symmetrical, triploblastic animals, and they are primarily distinguished by differences in early embryonic development, which shape how their body plans form. Protostomes include the vast majority of animal diversity, such as arthropods (insects, spiders, crustaceans) and mollusks (snails, clams, octopuses). Deuterostomes include chordates (which contain all vertebrates, including humans) and echinoderms (such as sea stars and sea urchins). Despite their differences, both groups share the same fundamental triploblastic, bilaterally symmetrical ancestor. The split between protostomes and deuterostomes represents one of the deepest and most important branches in animal evolution, leading to the vast diversity of body plans seen in animals today. Next, we will dive into protostomes and then come back to deuterostomes.
Protostomes vs. Deuterostomes: Blastopore
Kingdom Animalia: Eumetazoa: Bilateria: Nephrozoa: Protostomia vs. Deuterostomia
The key distinction comes early in development, during the formation of the blastopore, which is the first opening in the embryo. In protostomes, the blastopore becomes the mouth, while the anus forms later. In deuterostomes, this pattern is reversed: the blastopore becomes the anus, and the mouth forms second. This single developmental difference reflects deeper differences in how the embryo patterns its body axes and organ systems.
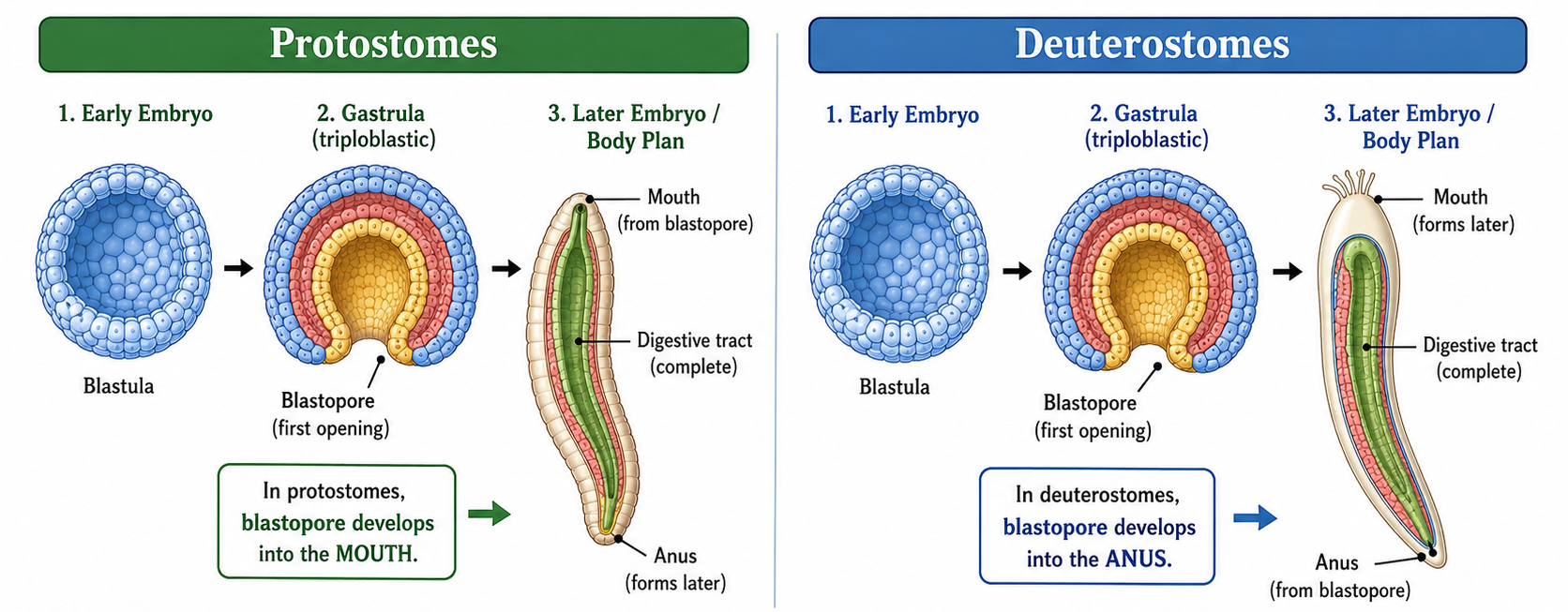
Figure 9. Blastopore fate in triploblastic animals. During gastrulation, the blastopore forms as the first opening of the embryo. In protostomes, the blastopore develops into the mouth and the anus forms later. In deuterostomes, the blastopore develops into the anus and the mouth forms later. This developmental difference marks one of the major evolutionary splits among bilaterally symmetrical triploblasts.
Protostomes and Deuterostomes: Early Embryonic Development
Kingdom Animalia: Eumetazoa: Bilateria: Nephrozoa: Protostomia vs. Deuterostomia
Protostomes develop using spiral and determinate cleavage. This means their early cells divide in a spiral pattern, and each cell’s future is set very early. If one of these early cells is removed, it usually cannot replace the others or form a full organism. As adults, protostomes are very diverse, and many groups like arthropods and annelids are segmented, with repeating body sections that can specialize for different jobs such as movement, feeding, or sensing. Deuterostomes develop using radial and indeterminate cleavage. Their early cells divide in a more organized, layered pattern, and the cells are not fixed in their fate early on. In many cases, early cells can still develop into a complete organism if separated. This flexibility helps support more complex body structures, including advanced organ systems found in animals like vertebrates.
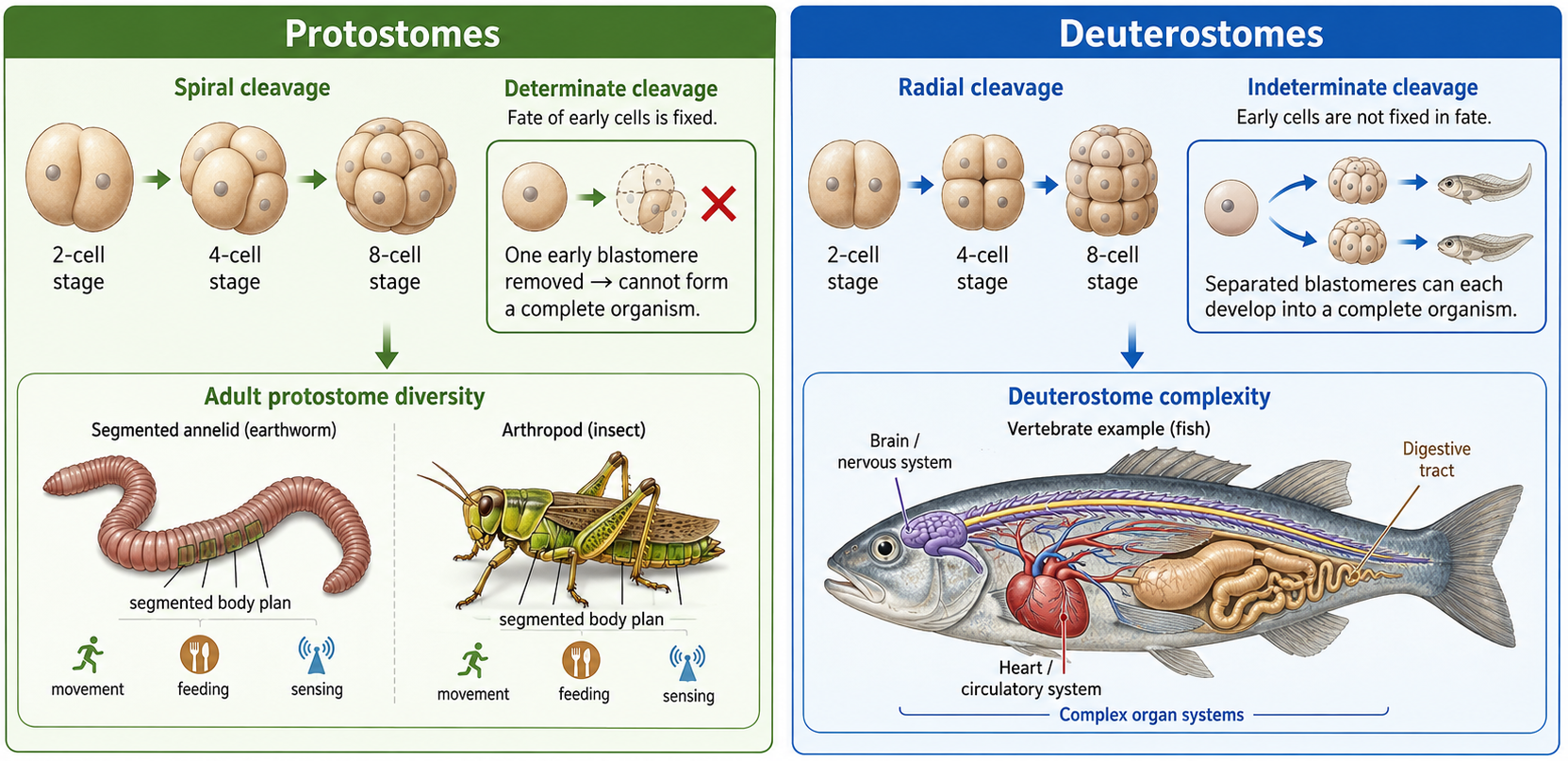
Figure 10. Comparison of protostome and deuterostome development. Protostomes typically show spiral, determinate cleavage, meaning early cell divisions are offset and cell fates are fixed early in development. Deuterostomes typically show radial, indeterminate cleavage, meaning early cells divide in aligned tiers and can retain developmental flexibility. These early developmental differences are associated with major differences in body organization, including segmentation in many protostomes and complex organ systems in deuterostomes such as vertebrates.
Protostomes and Deuterostomes: Coelom Development
Kingdom Animalia: Eumetazoa: Bilateria: Nephrozoa: Protostomia vs. Deuterostomia
The coelom forms in fundamentally different ways in protostomes and deuterostomes, and that difference reflects their deeper developmental split. In protostomes, the coelom forms when the mesoderm splits open internally, creating a fluid-filled body cavity, the coleom. Think of it as a block of tissue splitting open within the mesoderm to form a space. This cavity becomes the coelom, which later helps support organs and allows more complex body organization. In deuterostomes, the coelom forms when pockets of the endoderm pinch off and expand outward to form the coelom. Instead of splitting solid tissue, the body cavity forms from outpocketings of the digestive tract that separate and become internal cavities. Both processes produce a coelom, but they build it in opposite ways. Protostomes form it by splitting mesodermal tissue, while deuterostomes form it by budding off from the gut.
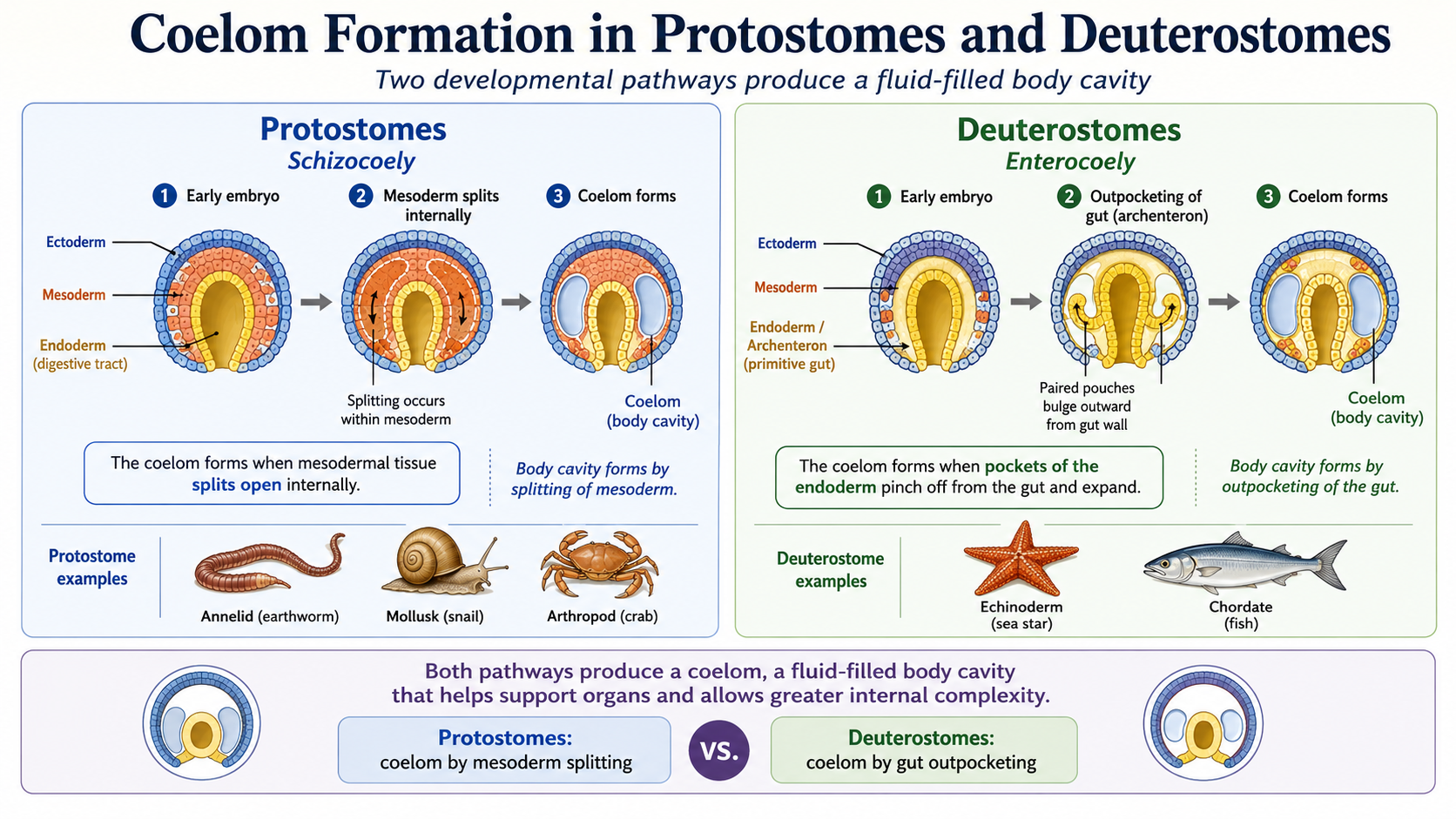
Figure 11. Coelom formation differs between the two major triploblastic lineages. In protostomes, the coelom forms by splitting within the mesoderm (schizocoely), whereas in deuterostomes, it forms when pockets of the endodermal gut pinch off and expand (enterocoely); both processes produce a fluid-filled body cavity that supports internal organs and greater body complexity.
Protostomes
Kingdom Animalia: Eumetazoa: Bilateria: Nephrozoa: Protostomia
Protostomes (Clade Protostomia) represent the largest and most diverse branch of the animal kingdom, containing the majority of all known animal species. Although many protostome groups are small, worm-like animals, the lineage also includes some of the most successful and recognizable animals on Earth. Among these are the arthropods, a group that includes insects, spiders, crustaceans, and their relatives. Arthropods alone account for most known animal species and have adapted to nearly every habitat, from the deepest oceans to deserts and forests. Despite their enormous diversity in form and lifestyle, all protostomes share a common pattern of embryonic development that distinguishes them from deuterostomes (see above). Protostomes are divided into two major evolutionary lineages: the Lophotrochozoa and the Ecdysozoa. While members of these groups can appear dramatically different from one another, they are distinguished by how they grow.
Lophotrochozoa
Kingdom Animalia: Eumetazoa: Bilateria: Nephrozoa: Protostomia: Lophotrochozoa
Lophotrochozoans generally grow continuously. As they increase in size, their tissues and supporting structures enlarge gradually. For example, worms grow by lengthening and widening their bodies, while mollusks such as clams and snails grow by expanding their soft tissues and adding new material to their shells. Growth in these animals is typically continuous rather than occurring in distinct stages.
Ecdysozoa
Kingdom Animalia: Eumetazoa: Bilateria: Nephrozoa: Protostomia: Ecdysozoa
Ecdysozoans, in contrast, grow by molting. These animals possess a tough external covering, such as an exoskeleton or cuticle, that cannot expand indefinitely. To increase in size, they must periodically shed this outer layer, enlarge their body, and then produce a new covering. Insects, spiders, crustaceans, and nematodes all grow in this way. As a result, growth in ecdysozoans occurs in a series of stages separated by molting events rather than as a continuous process.
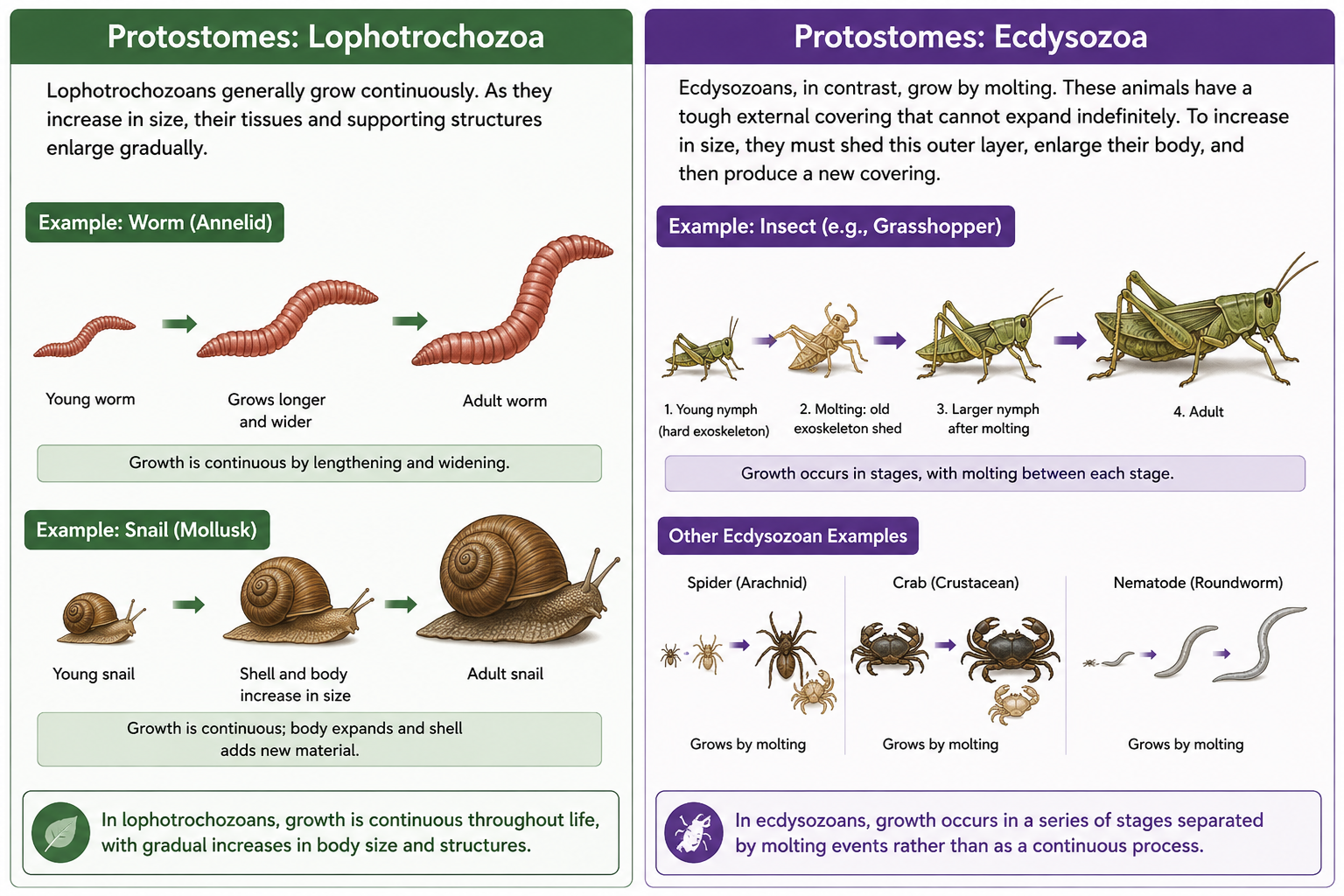
Figure 10. Growth Patterns in Protostomes. Protostomes are divided into two major clades that differ in how they grow. Lophotrochozoans grow continuously as their tissues and supporting structures gradually enlarge over time. Worms increase in length and width, while mollusks such as snails expand their soft tissues and add new material to their shells. In contrast, ecdysozoans grow through molting (ecdysis). Because their external cuticle or exoskeleton cannot expand indefinitely, they must periodically shed the old covering, enlarge their body, and produce a new one. As a result, growth in lophotrochozoans is gradual and continuous, whereas growth in ecdysozoans occurs in distinct stages separated by molting events. Examples of lophotrochozoans include annelid worms and mollusks, while ecdysozoans include insects, spiders, crustaceans, and nematodes.
Survey of Lophotrochozoa
Kingdom Animalia: Eumetazoa: Bilateria: Nephrozoa: Protostomia: Lophotrochozoa
Buckle your seatbelts. We are about to take a tour through the major protostome groups. We will begin with the Lophotrochozoa and then move on to the second major protostome lineage, the Ecdysozoa.
Phylum Rotifera (Rotifers)
Kingdom Animalia: Eumetazoa: Bilateria: Nephrozoa: Protostomia: Lophotrochozoa: Phylum Rotifera
Rotifers (Phylum Rotifera) are microscopic aquatic animals that are especially abundant in freshwater ponds, lakes, and streams, where they form an important part of the zooplankton community. Their most distinctive feature is the corona, a crown-like ring of beating cilia located at the anterior end of the body. The coordinated movement of these cilia creates water currents that draw suspended food particles, such as bacteria, algae, and organic debris, toward the mouth for ingestion. In addition to feeding, the corona also assists in locomotion, allowing many rotifers to swim through the water column. Despite their small size, rotifers possess specialized organ systems, including a complete digestive tract and a muscular pharynx called a mastax that contains hardened structures used to grind food. Their abundance and rapid reproduction make them important consumers of microorganisms and a vital food source for larger aquatic organisms.
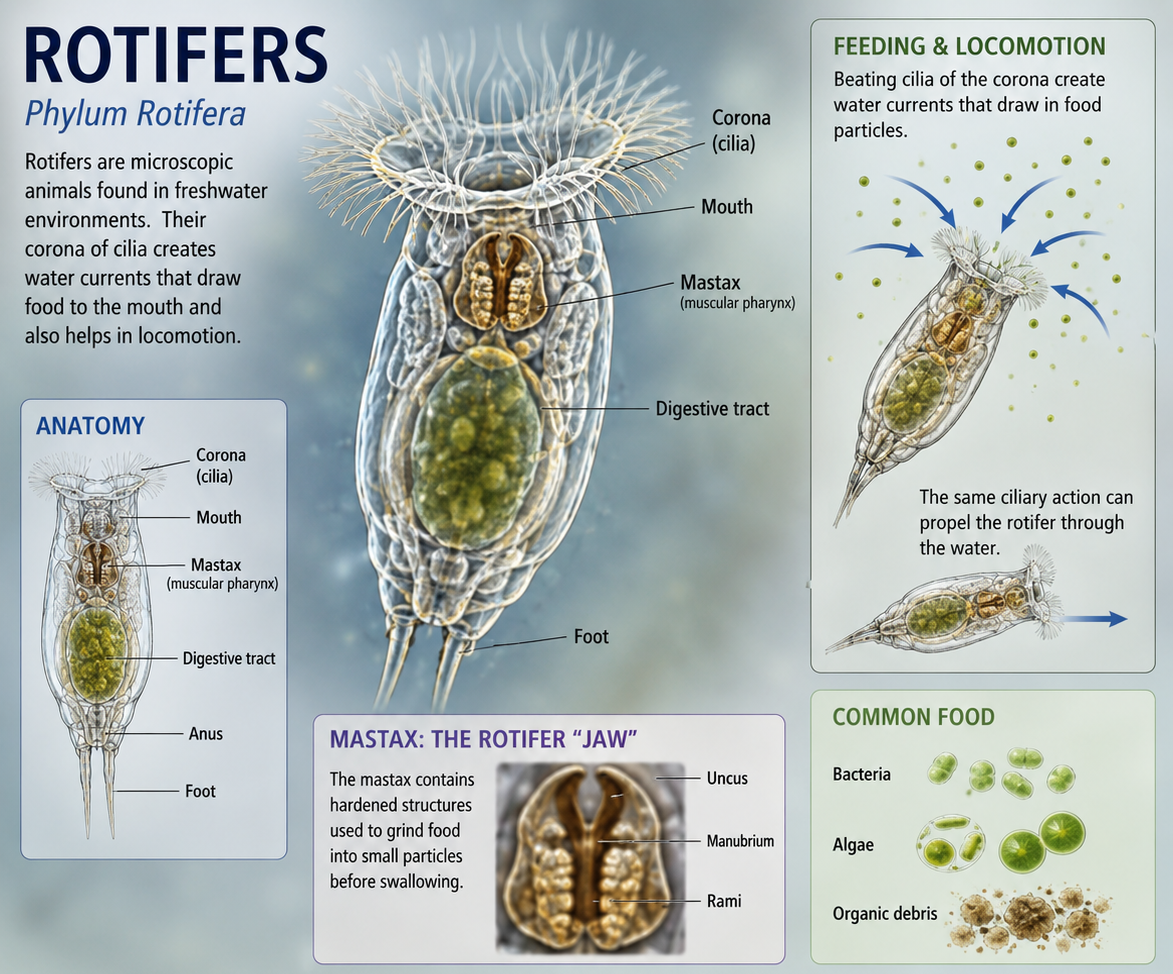
Figure 11. Rotifers (Phylum Rotifera). Rotifers are microscopic aquatic animals that are especially abundant in freshwater ponds, lakes, and streams, where they form an important component of the zooplankton community. Their most distinctive feature is the corona, a crown-like ring of beating cilia at the anterior end of the body that generates water currents for both feeding and locomotion. These currents draw suspended food particles, including bacteria, algae, and organic debris, toward the mouth. Rotifers possess a complete digestive tract and a specialized muscular pharynx called the mastax, which contains hardened structures used to grind food before ingestion. Despite their small size, rotifers play an important ecological role by consuming microorganisms and serving as a food source for larger aquatic organisms, helping transfer energy through freshwater food webs.
Phylum Platyhelminthes (Flatworms)
Kingdom Animalia: Eumetazoa: Bilateria: Nephrozoa: Protostomia: Lophotrochozoa: Phylum Platyhelminthes
The phylum Platyhelminthes includes the flatworms, a diverse group of bilaterally symmetrical animals characterized by their flattened bodies. Members of Platyhelminthes lack a coelom. Instead of having a fluid-filled body cavity between the digestive tract and the outer body wall, this space is filled with solid mesodermal tissue called parenchyma. This is confusing, because morpholigically Platyhelminthes resembles the condition seen in Acoelomorpha, an early-branching group of bilaterian animals. However, Acoelomorpha and Platyhelminthes are separate evolutionary lineages. Acoelomorpha branches near the base of bilaterian evolution, while Platyhelminthes belongs within the protostomes. Their shared lack of a coelom reflects a similar body plan, not membership in the same phylum.
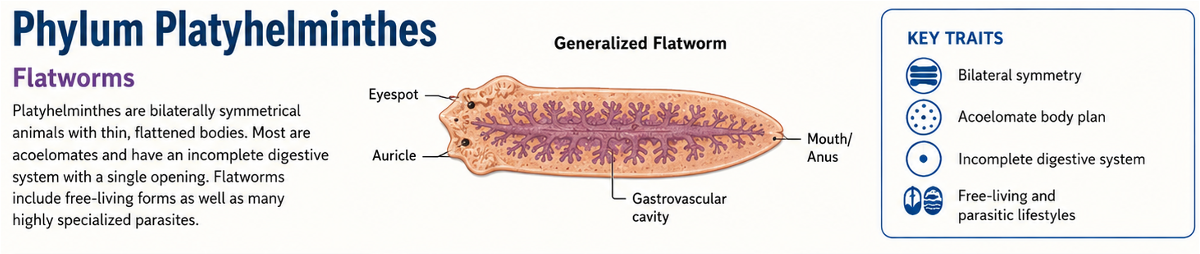
Figure 12. Phylum Platyhelminthes. Flatworms belong to Phylum Platyhelminthes and have flattened, bilaterally symmetrical bodies that lack a coelom. Instead of a fluid-filled body cavity, the space between the digestive tract and body wall is filled with solid mesodermal tissue called parenchyma. Although this resembles the condition seen in Acoelomorpha, the two groups are separate evolutionary lineages; Acoelomorpha branches near the base of Bilateria, while Platyhelminthes belongs within the protostomes.
Class Turbellaria (Turbellarians)
Kingdom Animalia: Eumetazoa: Bilateria: Nephrozoa: Protostomia: Lophotrochozoa: Phylum Platyhelminthes: Class Turbellaria
The turbellarians are primarily free-living flatworms that inhabit marine, freshwater, and moist terrestrial environments. Many species are especially common in coral reef ecosystems and other aquatic habitats. Turbellarians are typically predators or scavengers, feeding on protists, small invertebrates, and organic debris. They use a muscular, extendable pharynx to capture and ingest food. Unlike parasitic flatworms, turbellarians move actively through their environment and do not depend on a host for nutrition.

Figure 13. Class Turbellaria. Turbellarians are mostly free-living flatworms that inhabit marine, freshwater, and moist terrestrial environments. Unlike parasitic flatworms, they move actively through their habitats and feed as predators or scavengers on protists, small invertebrates, and organic debris rather than relying on a host for nutrition.
Class Cestoda (Tapeworms)
Kingdom Animalia: Eumetazoa: Bilateria: Nephrozoa: Protostomia: Lophotrochozoa: Phylum Platyhelminthes: Class Cestoda
The cestodes, commonly known as tapeworms, are highly specialized internal parasites that live in the digestive tracts of vertebrates. Because they inhabit an environment rich in pre-digested nutrients, they have completely lost their digestive system, including the mouth and gut. Instead, nutrients are absorbed directly through their body surface. Tapeworms consist of a head region, called a scolex, which attaches to the host's intestine, followed by a chain of reproductive segments. Their simplified anatomy reflects their adaptation to a parasitic lifestyle.
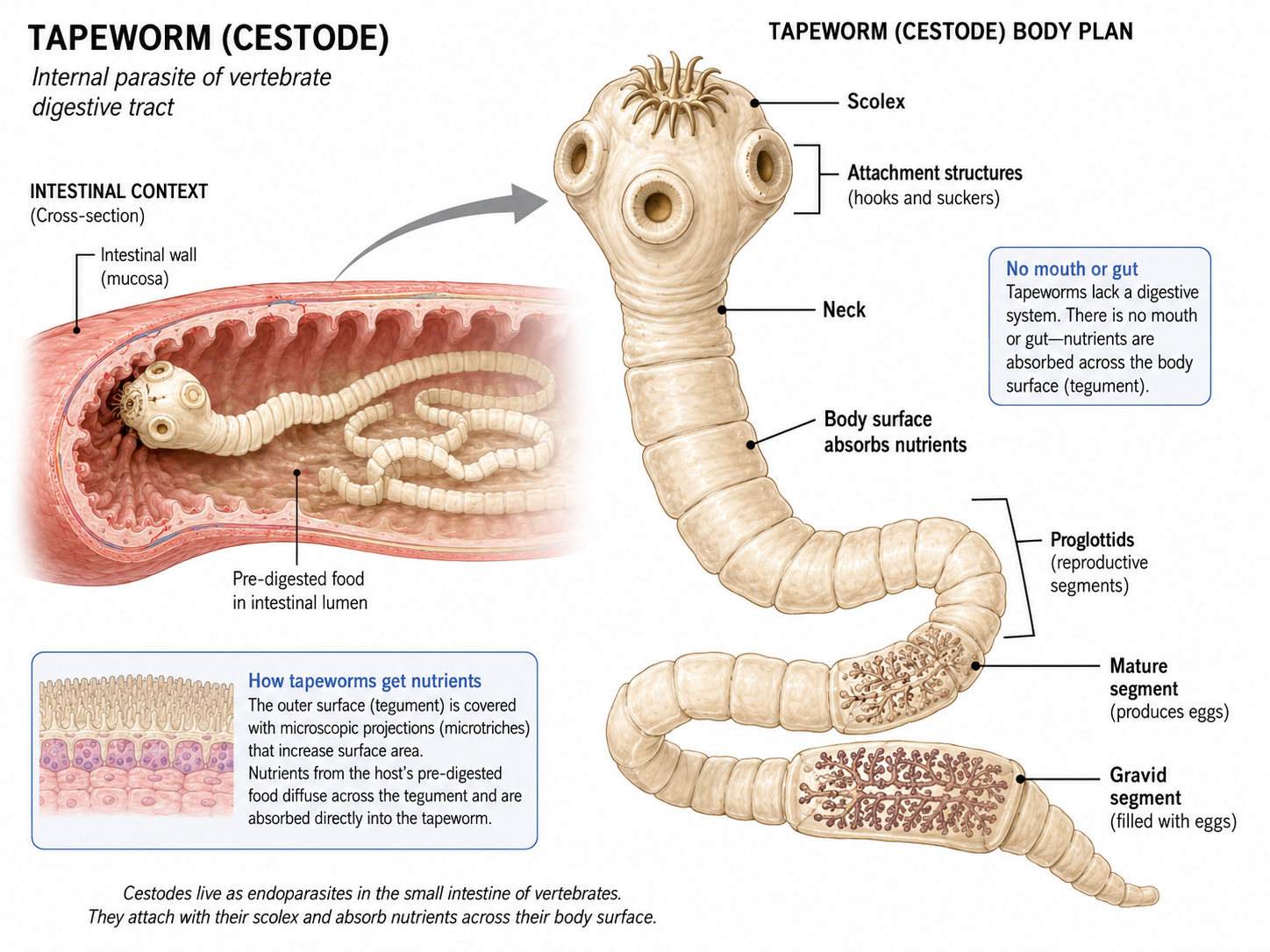
Figure 13. Class Cestoda. Cestode body plan. Tapeworms are highly specialized parasitic flatworms that live in the digestive tracts of vertebrates. The scolex anchors the worm to the intestinal wall, while the segmented body, made of reproductive proglottids, absorbs nutrients directly across the body surface because cestodes lack a mouth, gut, and digestive system.
Class Trematoda (Flukes)
Kingdom Animalia: Eumetazoa: Bilateria: Nephrozoa: Protostomia: Lophotrochozoa: Phylum Platyhelminthes: Class Trematoda
The trematodes, or flukes, are another major group of parasitic flatworms. Unlike tapeworms, trematodes possess a mouth and an incomplete digestive tract that allows them to ingest and digest host tissues, blood, or other bodily fluids. Many species have complex life cycles involving multiple hosts, often including snails as intermediate hosts and vertebrates as final hosts. Trematodes are responsible for several important diseases in humans and other animals and are among the most successful parasitic organisms on Earth.
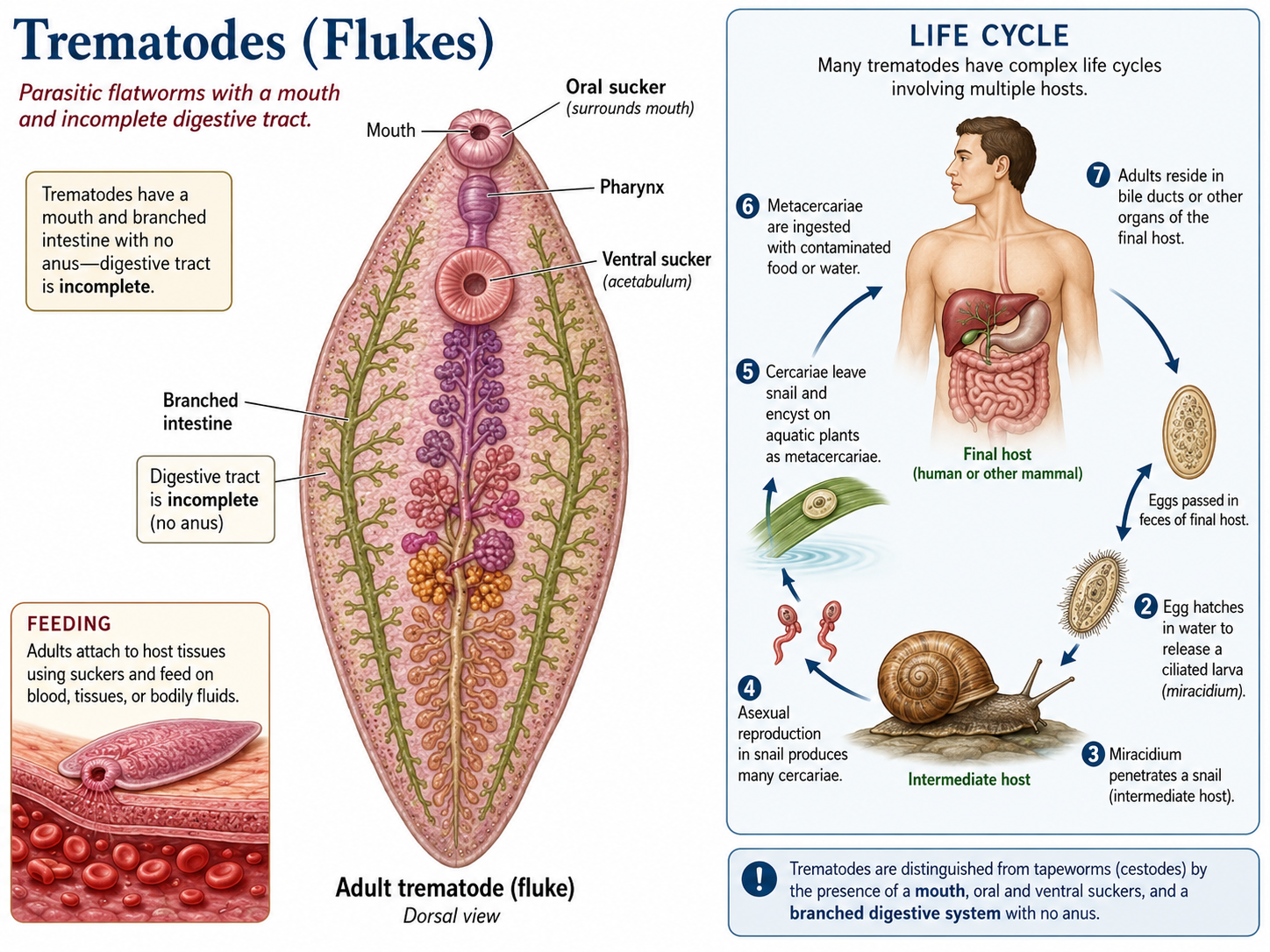
Figure 14. Class Trematoda. Trematodes are parasitic flatworms with a mouth, oral and ventral suckers, and an incomplete digestive tract with no anus. Unlike tapeworms, they ingest host tissues, blood, or bodily fluids, and many species have complex life cycles involving a snail intermediate host and a vertebrate final host.
Phylum Annelida (Segmented Worms)
Kingdom Animalia: Eumetazoa: Bilateria: Nephrozoa: Protostomia: Lophotrochozoa: Phylum Annelida
Annelids (Phylum Annelida) are segmented worms that include earthworms, many marine worms, and leeches. They live in a wide range of environments, including marine habitats, freshwater systems, and moist soils. Their defining feature is segmentation: the body is divided into repeated sections called segments. This body plan allows different regions of the body to specialize for movement, feeding, reproduction, and sensory functions. Most annelids have a true coelom, a fluid-filled body cavity lined by mesoderm. In many species, this fluid-filled cavity helps function as a hydrostatic skeleton. Circular and longitudinal muscles contract against the fluid pressure of the coelom, allowing the animal to crawl, burrow, or swim. Annelids also have a complete digestive tract, a closed circulatory system, and a centralized nervous system with paired nerve cords and segmental ganglia. Ecologically, annelids are important in many ecosystems. Earthworms mix and aerate soil, marine annelids help process organic material on the seafloor, and many annelids serve as prey for larger animals.
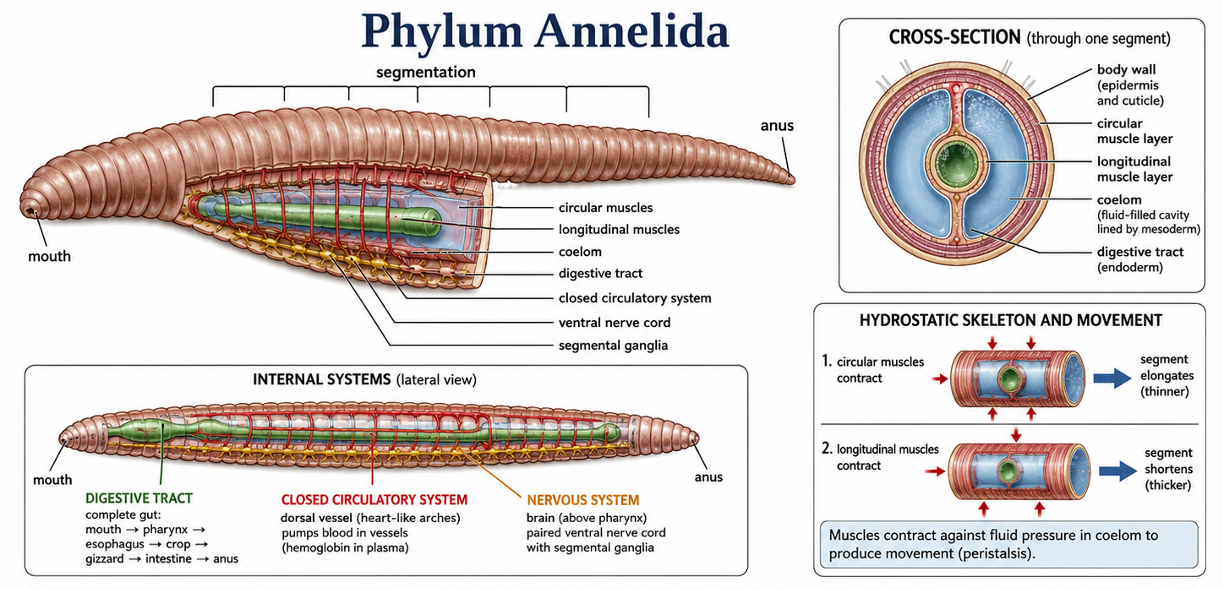
Figure 14. Phylum Annelida. Annelids are segmented worms with repeated body sections that allow different regions to specialize for movement, feeding, reproduction, and sensing. Most annelids have a true coelom that functions as a hydrostatic skeleton, allowing circular and longitudinal muscles to produce crawling, burrowing, or swimming movements. Their complete digestive tract, closed circulatory system, and centralized nervous system make them more complex than many earlier worm-like animals.
Class Polychaeta (Polychaete Worms)
Kingdom Animalia: Eumetazoa: Bilateria: Nephrozoa: Protostomia: Lophotrochozoa: Phylum Annelida: Class Polychaeta
Polychaete worms (Class Polychaeta) are mostly marine annelids and are often considered the group that most closely resembles early annelid body forms. The name Polychaeta means “many bristles,” which refers to the numerous chaetae found on their bodies. Chaetae are stiff, bristle-like structures made of chitin that help with movement, anchoring, and protection. In many polychaetes, the chaetae are located on paired, fleshy appendages called parapodia. These parapodia can function in crawling, swimming, burrowing, and gas exchange. Although polychaetes are not as familiar to most people as earthworms or leeches, they are extremely important in marine food webs and seafloor ecosystems.
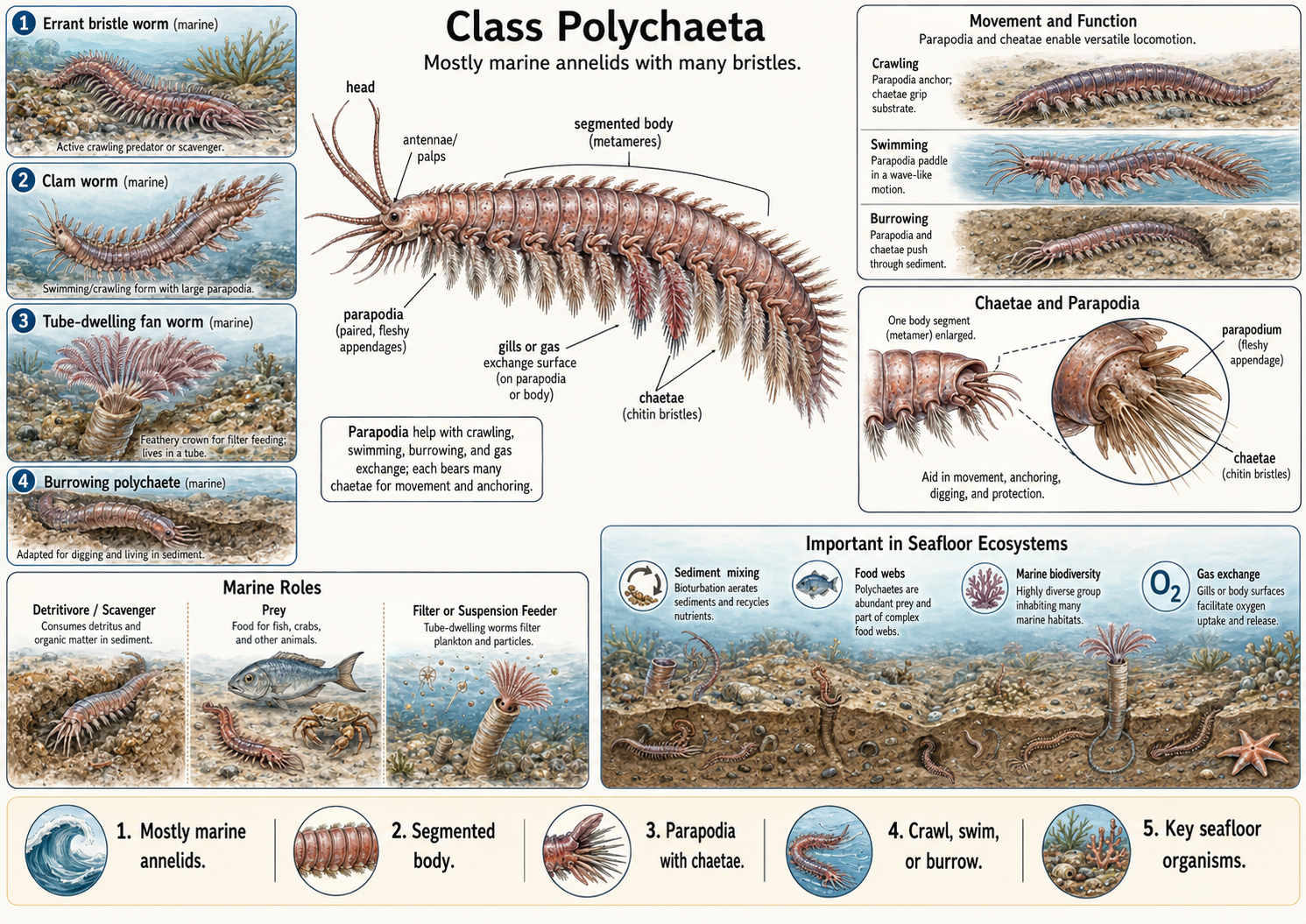
Figure 15. Class Polychaeta. Polychaete worms are mostly marine annelids characterized by segmented bodies, numerous chaetae, and paired fleshy appendages called parapodia. The chaetae and parapodia help with crawling, swimming, burrowing, anchoring, and, in some species, gas exchange. Polychaetes include active errant worms, burrowing forms, and tube-dwelling suspension feeders. This figure shows their basic body plan, the structure and function of chaetae and parapodia, different modes of locomotion, and the important ecological roles polychaetes play in marine food webs and seafloor ecosystems.
Class Oligochaeta (Oligochaete worms, i.e. earthworms)
Kingdom Animalia: Eumetazoa: Bilateria: Nephrozoa: Protostomia: Lophotrochozoa: Phylum Annelida: Class Oligochaeta
Oligochaete worms (Class Oligochaeta) are annelids that include earthworms and many freshwater worms. The name Oligochaeta means “few bristles,” which reflects the reduced number of chaetae compared with polychaetes. Unlike many polychaetes, oligochaetes lack parapodia, but they still have small chaetae that help them grip the soil or sediment during movement. Earthworms are the best-known members of this group. As they burrow and feed, they break down organic material, mix soil layers, and improve soil structure.
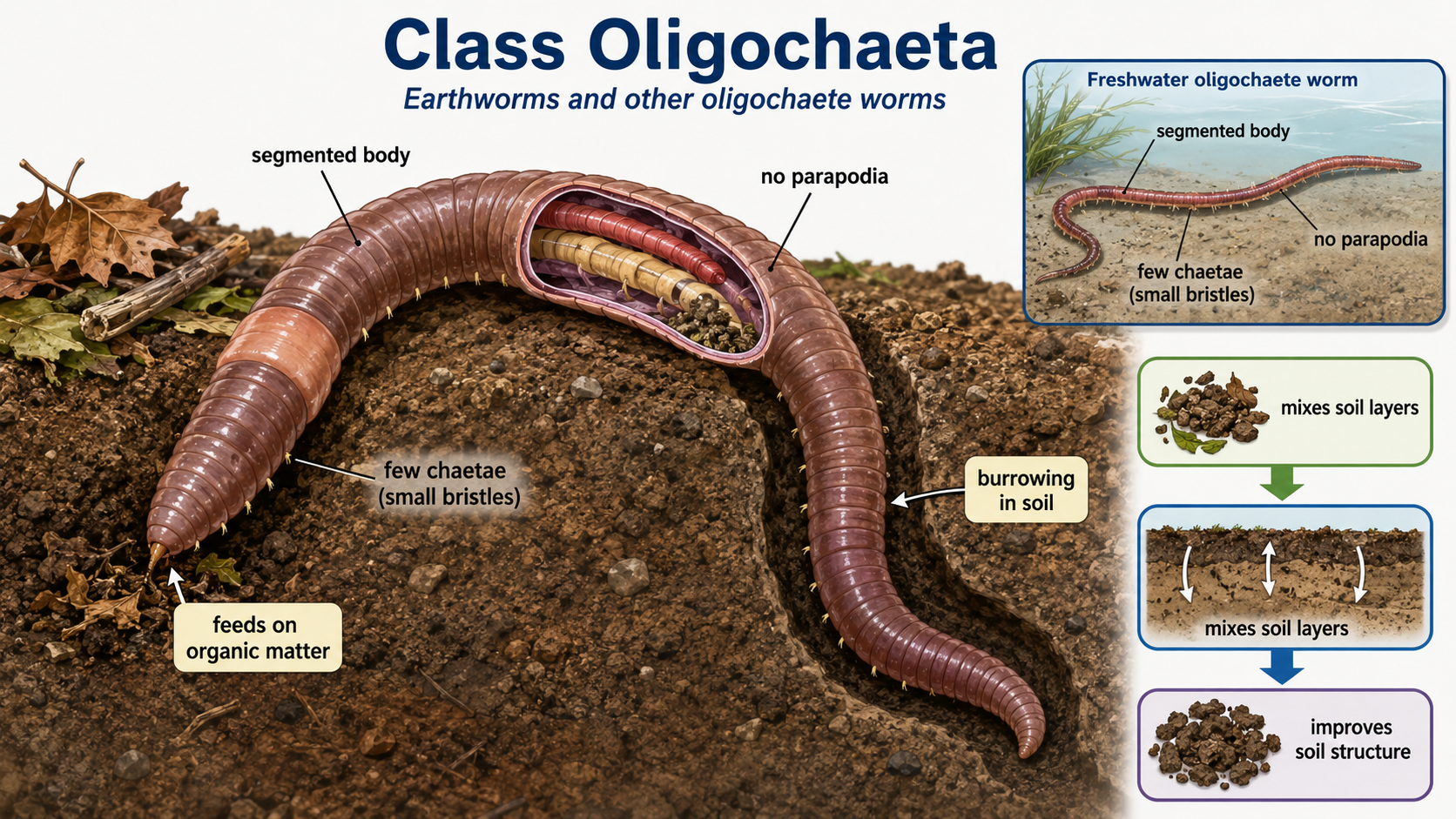
Figure 16. Class Oligochaeta. Oligochaete worms, including earthworms, are annelids with segmented bodies, few chaetae, and no parapodia. Their small bristles help them move through soil or sediment, and their burrowing and feeding activities break down organic matter, mix soil layers, and improve soil structure.
Class Hirudinea (Leeches)
Kingdom Animalia: Eumetazoa: Bilateria: Nephrozoa: Protostomia: Lophotrochozoa: Phylum Annelida: Class Hirudinea
Class Hirudinea is the annelid group that includes leeches. Leeches have bodies that are flattened and segmented, but their external segmentation is modified compared with earthworms and polychaetes. Most leeches lack parapodia and chaetae. Instead, they move using muscular body contractions and suckers located at the anterior and posterior ends of the body. Some leeches feed on blood, but many are predators or scavengers that consume small invertebrates. Although leeches are often known for their blood-feeding behavior, they are a diverse group with important ecological roles in freshwater and moist terrestrial habitats.

Figure 17. Class Hirundinea. Leeches are annelids in Class Hirudinea with flattened, segmented bodies and anterior and posterior suckers used for attachment, feeding, and movement. Although some species feed on blood, many leeches are predators or scavengers, making them important members of freshwater and moist terrestrial ecosystems.
Phylum Mollusca (Mollusks)
Kingdom Animalia: Eumetazoa: Bilateria: Nephrozoa: Protostomia: Lophotrochozoa: Phylum Mollusca
Mollusks (Phylum Mollusca) are a diverse group of soft-bodied animals that include snails, slugs, clams, oysters, squid, octopuses, and chitons. They live in many environments, including marine habitats, freshwater systems, and moist terrestrial habitats. Although mollusks vary greatly in body shape, most share three major body regions: a muscular foot, a visceral mass, and a mantle. The muscular foot is a major part of the mollusk body plan, but its function varies among groups. In snails and slugs, it is broad and flat for crawling. In clams, it is shaped for burrowing into sand or mud. In some mollusks, it helps with attachment, while in cephalopods such as squid and octopuses, it is modified into arms and tentacles used for capturing prey and movement. The visceral mass contains most of the internal organs, including those used for digestion, reproduction, circulation, and excretion. The mantle is a sheet of tissue that covers and protects this region. In many mollusks, the mantle secretes a calcium carbonate shell that provides protection and support. In groups such as slugs and octopuses, the shell is reduced or absent, but the mantle remains an important body structure.
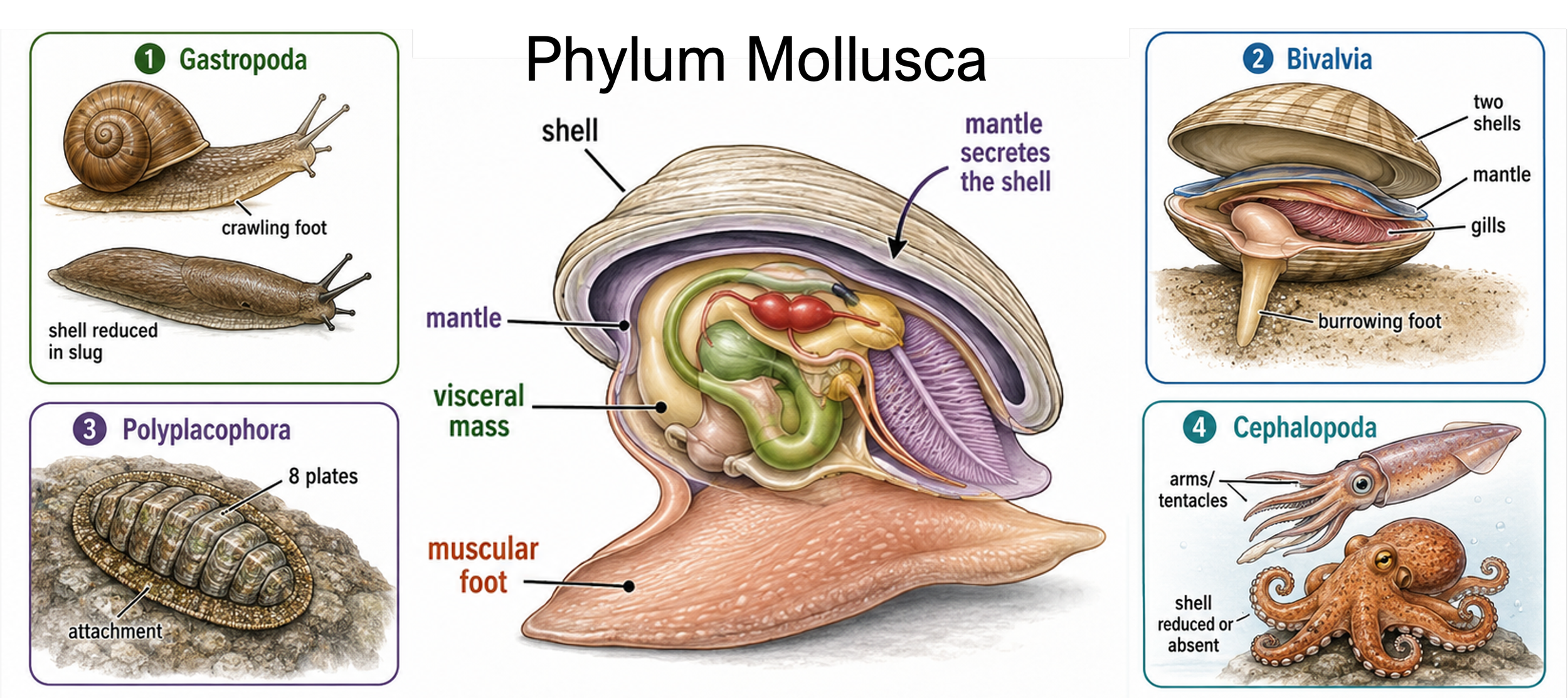
Figure 18. Mollusk body plan and diversity. Mollusks (Phylum Mollusca) share three major body regions: a muscular foot, a visceral mass, and a mantle. The foot is modified for crawling, burrowing, attachment, or grasping; the visceral mass contains the internal organs; and the mantle protects the body and often secretes a shell. Major mollusk groups include gastropods, bivalves, chitons, and cephalopods, which occupy marine, freshwater, and moist terrestrial habitats.
Class Gastropoda (Gastropods)
Kingdom Animalia: Eumetazoa: Bilateria: Nephrozoa: Protostomia: Lophotrochozoa: Phylum Mollusca: Class Gastropoda
Gastropods are the largest and most diverse group of mollusks in Class Gastropoda. This group includes snails, slugs, limpets, conchs, nudibranchs, and many other familiar mollusks. Most gastropods have a broad muscular foot used for crawling, and many have a single coiled shell for protection. However, in some groups, such as slugs and nudibranchs, the shell has been reduced or lost. Most gastropods feed using a radula, a ribbon-like scraping organ covered with tiny teeth. Herbivorous gastropods use the radula to scrape algae or plant material from surfaces, while predatory gastropods may use it to drill into shells or tear apart prey. This flexible feeding structure has helped gastropods adapt to many different diets and habitats. Gastropods live in marine, freshwater, and terrestrial environments. Marine snails and limpets often graze on algae along rocks and reefs, freshwater snails feed in ponds and streams, and land snails and slugs move through moist soils and vegetation. Their muscular foot, radula, and protective shell have made gastropods one of the most successful mollusk groups.

Figure 19. Class Gastropoda. Gastropods are the largest and most diverse class of mollusks. Most move with a broad muscular foot and feed with a radula, a ribbon-like scraping structure used in both herbivorous and predatory feeding. Many gastropods have a single coiled shell, although the shell is reduced or absent in groups such as slugs and nudibranchs. This figure shows the general gastropod body plan, examples of major gastropod forms, different uses of the radula, and the presence of gastropods in marine, freshwater, and terrestrial environments.
Class Polyplacophora (Chitons)
Kingdom Animalia: Eumetazoa: Bilateria: Nephrozoa: Protostomia: Lophotrochozoa: Phylum Mollusca: Class Polyplacophora
Chitons are a strange but important group of mollusks in Class Polyplacophora. Like gastropods, they have a broad muscular foot used for crawling and a radula used for feeding. However, chitons are easy to recognize because their backs are covered by eight overlapping shell plates. These plates act like armor, protecting the soft body while still allowing the animal to bend and grip uneven rock surfaces. This differs from snails, which usually have a single coiled shell. Most chitons live in marine environments, especially along rocky shorelines. They use their strong muscular foot to cling tightly to rocks, helping them resist waves and avoid predators. Many chitons are grazers that scrape algae and other small organisms from rock surfaces using a radula that is often strengthened with hard minerals. Their flattened shape, powerful foot, and flexible armor make chitons well adapted to life on exposed marine surfaces. Ecologically, they help control algal growth and serve as prey for sea stars, crabs, fish, and shorebirds.

Figure 20. Class Polyplacophora. Chiton body form and ecology. Chitons are marine mollusks in Class Polyplacophora that are adapted for life on rocky shorelines. Their backs are protected by eight overlapping shell plates, which act as flexible armor while allowing the body to bend against uneven rock surfaces. The broad muscular foot helps the animal crawl and cling tightly to rocks, and the radula scrapes algae and other small organisms from the surface. Chitons differ from snails, which usually have a single coiled shell, and they serve as prey for sea stars, crabs, fish, and shorebirds.
Class Bivalvia (Bivalves)
Kingdom Animalia: Eumetazoa: Bilateria: Nephrozoa: Protostomia: Lophotrochozoa: Phylum Mollusca: Class Bivalvia
Bivalves are mollusks in Class Bivalvia, a group that includes clams, oysters, mussels, and scallops. Their name comes from their body structure: they have two shells, or valves, that are attached by a hinge. These shells protect the soft body inside and can be opened or closed by strong muscles. Unlike many other mollusks, bivalves do not have a radula. Instead, they are suspension feeders, meaning they filter tiny food particles from the water. Clams usually live buried in sand or mud. They extend tube-like structures called siphons up into the water. One siphon draws water into the body, where food particles are trapped by the gills, and the other siphon releases filtered water back out. Oysters and mussels also filter feed, but they usually attach themselves to hard surfaces such as rocks, shells, docks, or reefs. By filtering large amounts of water, these animals help remove suspended particles and improve water clarity. Scallops are different from many other bivalves because they are more mobile. Instead of staying buried or permanently attached, scallops can swim short distances by rapidly opening and closing their shells. This movement pushes water out and allows the scallop to escape some predators. Although bivalves may look simple, they play major ecological roles as filter feeders, habitat builders, and prey for many marine and freshwater animals.
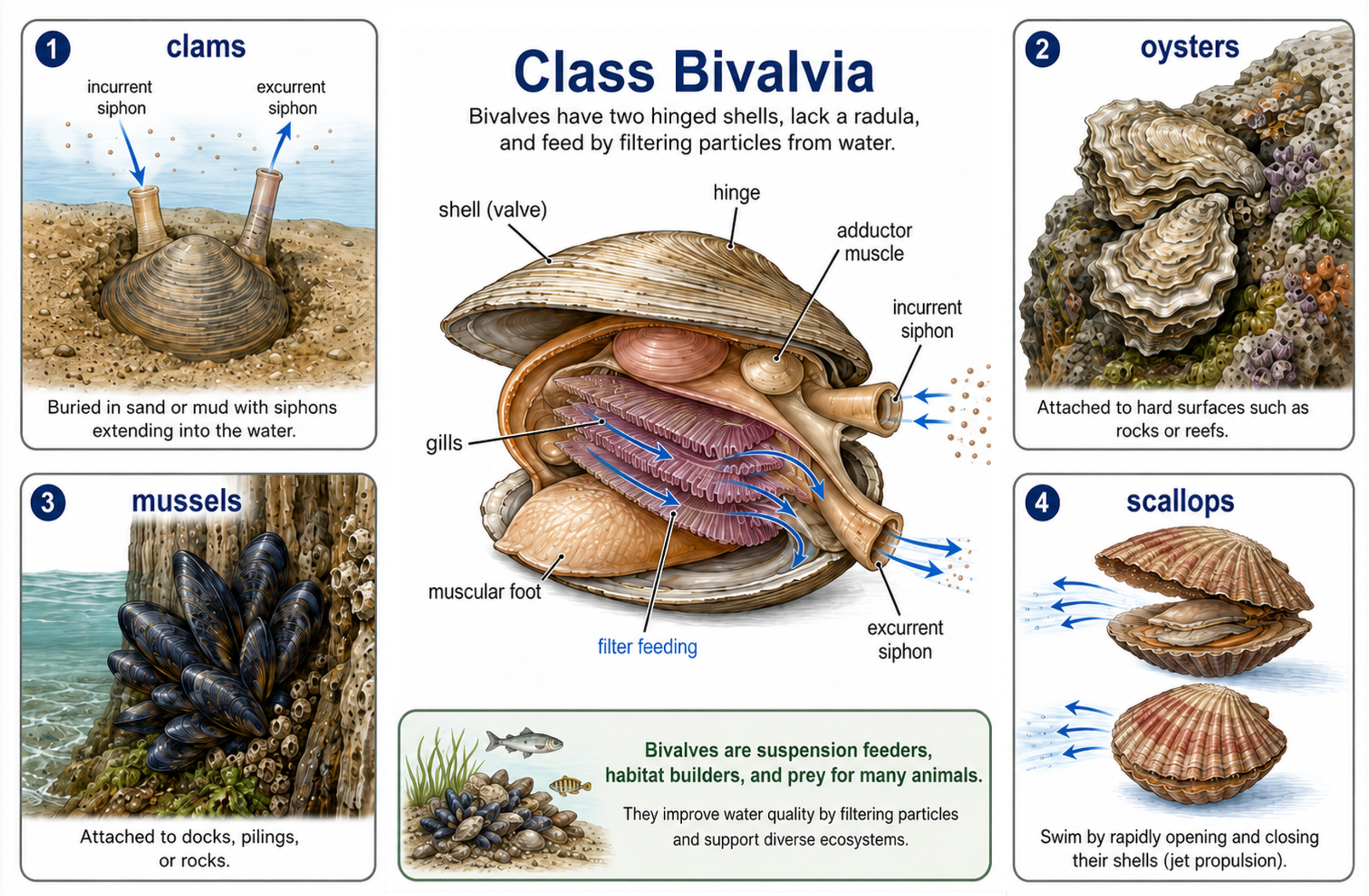
Figure 21. Class Bivalvia anatomy and ecological roles. Bivalves, including clams, oysters, mussels, and scallops, are mollusks with two hinged shells and no radula. They feed by drawing water through an incurrent siphon, filtering food particles across the gills, and releasing filtered water through an excurrent siphon. Different bivalves live in different ways: clams often burrow in sediment, oysters and mussels attach to hard surfaces, and scallops can swim by rapidly opening and closing their shells. As suspension feeders, bivalves improve water clarity, build habitat, and serve as prey for many aquatic animals.
Class Cephalopoda (Cephalopods)
Kingdom Animalia: Eumetazoa: Bilateria: Nephrozoa: Protostomia: Lophotrochozoa: Phylum Mollusca: Class Cephalopoda
Cephalopods are mollusks in Class Cephalopoda, a group that includes nautiluses, cuttlefish, squid, and octopuses. They may not look much like clams or snails, but they share the same basic mollusk ancestry. Their name means “head-foot,” which refers to the way the mollusk foot has been modified into arms and tentacles around the head. These appendages are used for grasping prey, crawling, swimming, and exploring the environment. Cephalopods have a well-developed head, large eyes, and a strong beak used to bite and tear food. In octopuses, the beak is the main hard structure in the body, which allows them to squeeze through very small spaces. Squid and cuttlefish have internal shell-like supports, while the nautilus still has an external chambered shell. Most cephalopods are active predators that feed on fish, crustaceans, and other mollusks. Cephalopods are the most intelligent protostomes and have complex nervous systems, advanced behaviors, and strong problem-solving abilities. Octopuses can open containers, escape enclosures, use camouflage, and learn from experience. In this sense, cephalopods are like the “great apes” of the protostome world: highly active, behaviorally complex animals that evolved intelligence along a very different path from vertebrates.

Figure 22. Class Cephalopoda. Cephalopods are active marine mollusks that include nautiluses, cuttlefish, squid, and octopuses. Their molluscan foot has been modified into arms and tentacles used for grasping prey, movement, and exploration. Most cephalopods have large eyes, a well-developed nervous system, and a strong beak used to bite and tear food. The nautilus retains an external chambered shell, squid and cuttlefish have internal shell-like supports, and octopuses have a greatly reduced or absent shell. Cephalopods are active predators and are the most intelligent protostomes, showing complex behaviors such as camouflage, problem solving, learning, and object manipulation.
Survey of Ecdysozoa
Kingdom Animalia: Eumetazoa: Bilateria: Nephrozoa: Protostomia: Ecdysozoa
We now turn from the Lophotrochozoa to the second major lineage of protostomes, the Ecdysozoa.
Phylum Nematoda (Roundworms)
Kingdom Animalia: Eumetazoa: Bilateria: Nephrozoa: Protostomia: Ecdysozoa: Phylum Nematoda
Nematodes (Phylum Nematoda), commonly known as roundworms, are among the most abundant and widespread animals on Earth. They occur in nearly every habitat, including marine environments, freshwater systems, soils, deserts, polar regions, mountains, deep-sea trenches, and even underground environments. Nematodes outnumber all other animals in terms of total numbers on Earth. They are especially abundant in sediments and soils, where they play important roles in nutrient cycling, decomposition, and food webs. Nematodes occupy a wide range of ecological roles. Many species are free-living and feed on bacteria, fungi, algae, or other small organisms, while others are predators. Many species are also parasitic, living within plants, animals, and humans. These parasitic forms can cause significant diseases in crops, livestock, wildlife, and people. Because of their extraordinary abundance, ecological diversity, and presence at multiple trophic levels, nematodes are among the most important animals in terrestrial and aquatic ecosystems.
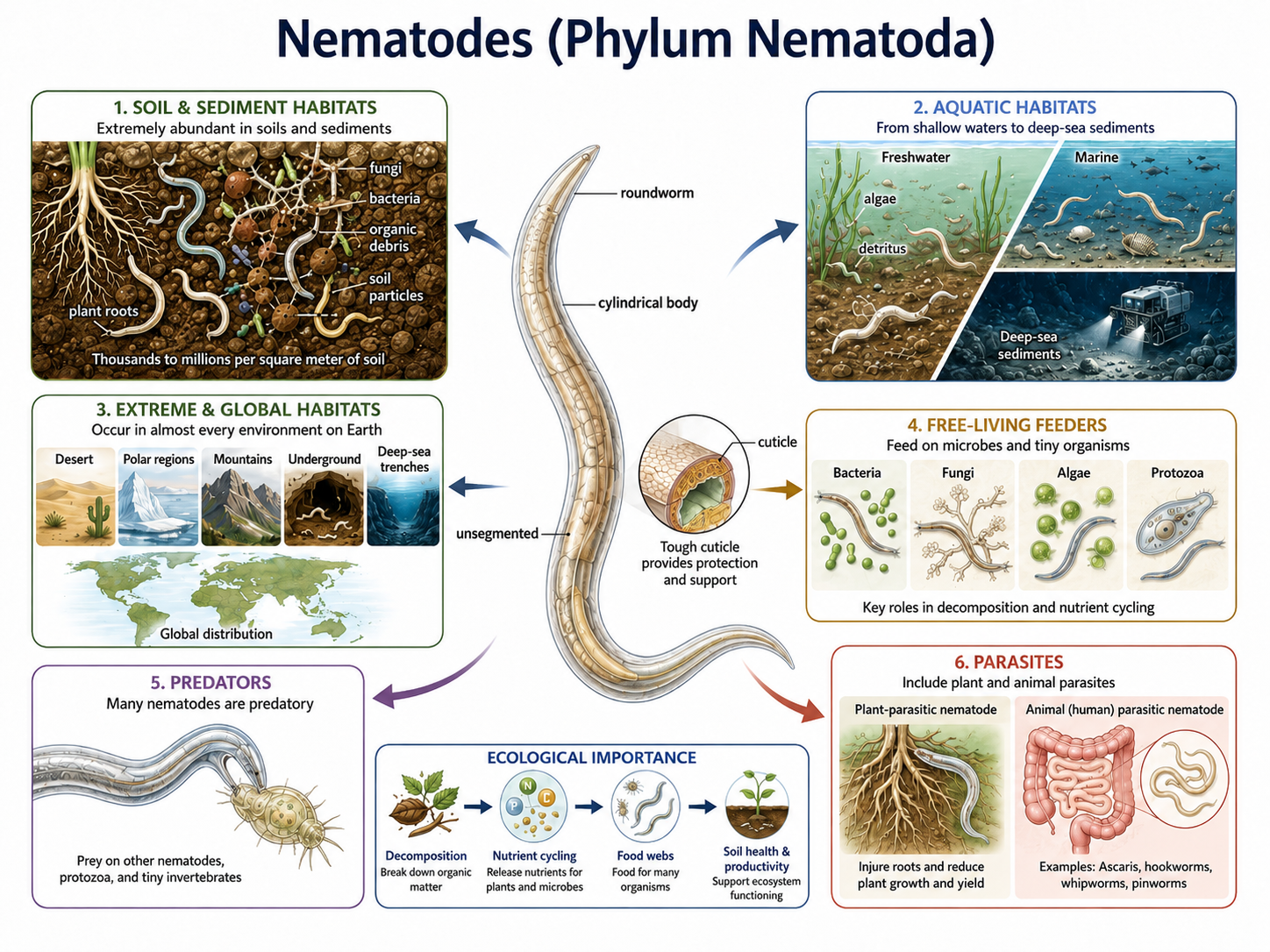
Figure 23. Nematodes (Phylum Nematoda) are abundant roundworms with diverse ecological roles. Nematodes have slender, cylindrical, unsegmented bodies covered by a tough cuticle. They occur in soils, sediments, freshwater, marine habitats, and extreme environments around the world. Many free-living species feed on bacteria, fungi, algae, protozoa, or other small organisms, contributing to decomposition, nutrient cycling, soil health, and food webs. Other nematodes are predators or parasites of plants and animals.
Phylum Tardigrada (Water Bears)
Kingdom Animalia: Eumetazoa: Bilateria: Nephrozoa: Protostomia: Ecdysozoa: Phylum Tardigrada
Tardigrades (Phylum Tardigrada), commonly known as water bears, are microscopic animals that live in thin films of water in mosses, lichens, soils, freshwater habitats, and marine sediments. They have a segmented body with four pairs of short legs, giving them eight legs total. Their name means “slow walker,” and they are called water bears because their slow, lumbering movement resembles a bear’s gait. Tardigrades are famous for surviving extreme conditions. When the environment becomes unfavorable, they can enter a dormant state called cryptobiosis, drying out and curling into a compact form called a tun. In this state, their metabolism drops to nearly undetectable levels, allowing some species to survive extreme cold, intense heat, high radiation, dehydration, high pressure, and even short-term exposure to the vacuum of space. Their resilience comes partly from protective molecules that stabilize their cells and DNA during stress. However, tardigrades are not indestructible; they are much more vulnerable when active and hydrated. Most feed on plant cells, algae, bacteria, or tiny invertebrates using sharp mouthparts called stylets to pierce cells and suck out their contents.

Figure 24. Phylum Tardigarda. Tardigrades (Phylum Tardigrada), commonly known as water bears, are microscopic eight-legged animals that live in thin films of water in mosses, lichens, soils, freshwater habitats, and marine sediments. They are famous for surviving extreme conditions by entering cryptobiosis, a dormant tun state in which their metabolism nearly stops, though they are not indestructible and usually feed on plant cells, algae, bacteria, or tiny invertebrates using sharp stylets.
Phylum Onychophora (Velvet Worms)
Kingdom Animalia: Eumetazoa: Bilateria: Nephrozoa: Protostomia: Ecdysozoa: Phylum Onychophora
Velvet worms (Phylum Onychophora) are soft-bodied animals that look like a mix between a worm, caterpillar, and slug. Their name means “claw bearers,” referring to the tiny claws at the ends of their many short legs. Although they resemble caterpillars, they are not insects. Most velvet worms live in moist, dark habitats, especially in tropical and Southern Hemisphere regions. They have small eyes, antennae, soft segmented bodies, many paired legs, and specialized slime glands near the mouth. When hunting, they shoot sticky slime at insects and other small invertebrates, trapping prey before feeding. Velvet worms are important evolutionarily because they combine worm-like softness with arthropod-like traits such as clawed legs. Many species also have unusual reproduction, including live birth and long-term sperm storage. Their strange body plan, slime-based hunting, and evolutionary position make them one of the most fascinating animal groups.

Figure 23. Phylum Onychophora. Velvet worms are soft-bodied animals in Phylum Onychophora that live in moist, dark habitats and move on many short legs with tiny claws. Their antennae, slime glands, and sticky slime-based hunting distinguish them from insects and highlight their evolutionary link between worm-like body forms and arthropod-like traits.
Phylum Arthropoda (Arthropods)
Kingdom Animalia: Eumetazoa: Bilateria: Nephrozoa: Protostomia: Ecdysozoa: Phylum Arthropoda
Arthropods (Phylum Arthropoda) are invertebrate animals with a hard external skeleton, a segmented body, and jointed appendages. The name Arthropoda comes from Greek words meaning “jointed foot” or “jointed leg.” This phylum includes insects, spiders, scorpions, crustaceans, centipedes, millipedes, and many other groups. The arthropod body is covered by a tough outer cuticle made mostly of chitin. This cuticle functions as an exoskeleton, protecting the body, preventing water loss, and providing attachment points for muscles. In crustaceans, such as crabs and lobsters, the exoskeleton is often strengthened with calcium carbonate, making it especially hard. However, because the exoskeleton does not grow with the animal, arthropods must periodically shed it through a process called molting. After molting, the new exoskeleton is soft for a short time, leaving the animal vulnerable until it hardens. A major reason arthropods are so successful is their highly flexible body plan. Their bodies are built from repeated segments, and many segments bear paired appendages. Over evolutionary time, these appendages have been modified for walking, swimming, feeding, sensing, mating, defense, and flight. This “Swiss Army knife” body plan allowed arthropods to adapt to an enormous variety of ecological roles. Arthropods have the highest species diversity of all animal phylums, with more than a million described species and many more still undescribed. They are found in nearly every environment, including oceans, freshwater systems, forests, deserts, soils, caves, and even human homes. Along with amniotes, arthropods are one of the few animal groups to become highly successful in dry terrestrial environments. Their success is largely due to their waterproof exoskeleton, efficient movement, specialized appendages and in insects, the evolution of flight. Arthropods range dramatically in size, from microscopic planktonic forms to large crabs with leg spans of several meters. Despite this diversity, they are united by the same core traits: a segmented body, jointed appendages, an exoskeleton, and growth by molting.

Figure 24. Phylum Arthropoda. Arthropods share a segmented body, paired jointed appendages, and a hard chitinous cuticle that functions as an exoskeleton for protection, water-loss prevention, and muscle attachment. The surrounding examples illustrate the diversity of the phylum, including insects, arachnids, crustaceans, centipedes, and millipedes, while the molting inset shows how arthropods shed and replace the exoskeleton as they grow.
Subphylum Myriapoda (Millipedes and Centipedes)
Kingdom Animalia: Eumetazoa: Bilateria: Nephrozoa: Protostomia: Ecdysozoa: Phylum Arthropoda: Subphylum Myriapoda
Myriapods (Subphylum Myriapoda) are arthropods that include millipedes and centipedes. Their name means “many feet,” which fits their long, segmented bodies and numerous walking legs. Although millipedes and centipedes may look similar at first, they differ strongly in body structure, feeding behavior, and lifestyle. Millipedes are mostly detritivores, meaning they feed on decaying leaves, dead plant material, and other organic matter. By breaking down plant debris, they help recycle nutrients back into the soil. Millipedes usually have two pairs of legs per body segment, giving them a slow, steady walking style. They are generally not predators and often defend themselves by curling into a coil or releasing unpleasant chemical secretions. Centipedes, in contrast, are active predators. They have one pair of legs per body segment and are usually faster-moving than millipedes. Their first pair of appendages is modified into venomous claws called forcipules, which they use to capture and subdue prey such as insects and other small animals. While millipedes are slow-moving decomposers with two leg pairs per segment, centipedes are fast-moving predators with one leg pair per segment.
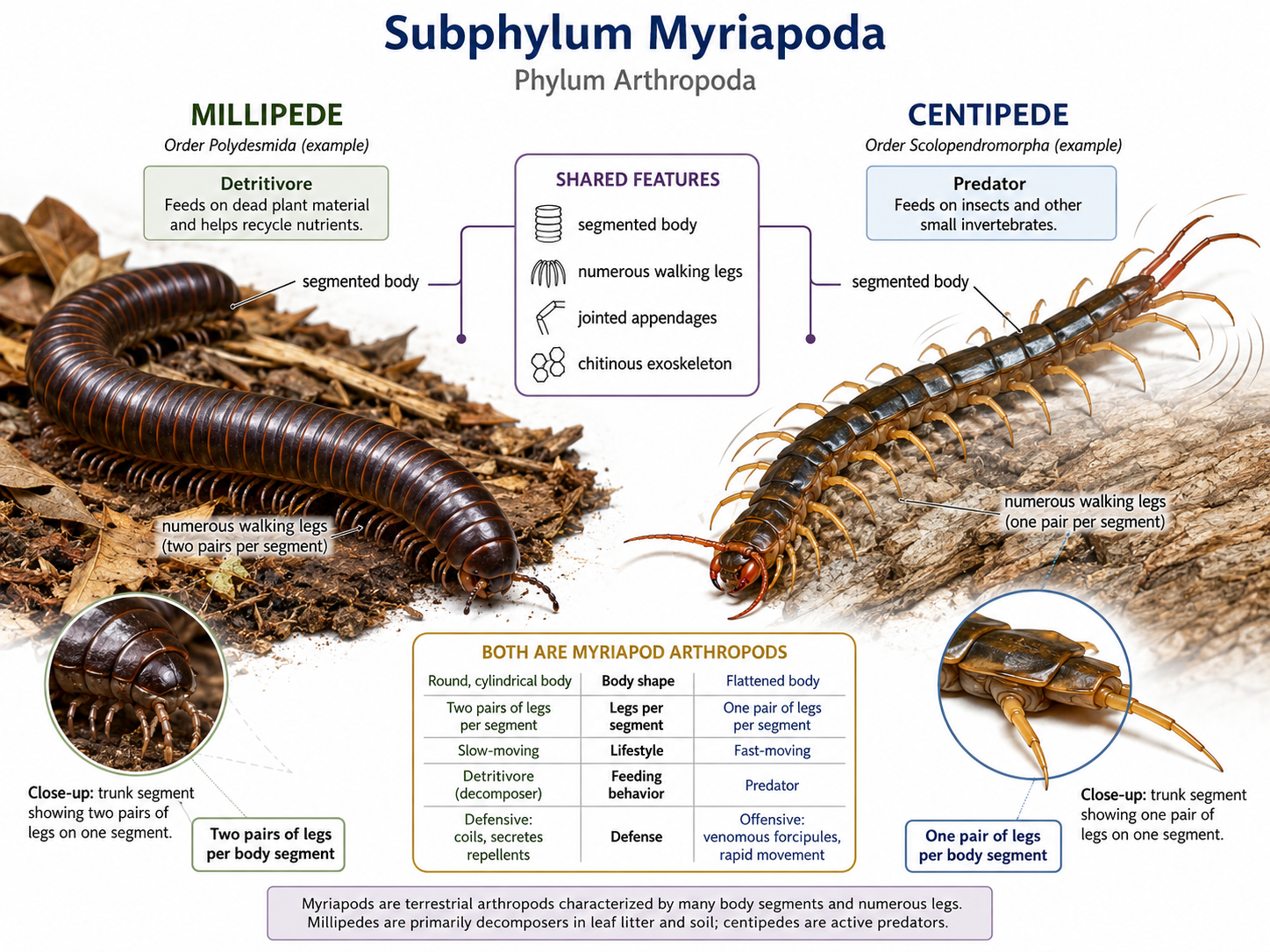
Figure 25. Subphylum Myriapoda: millipedes and centipedes. Millipedes and centipedes are myriapod arthropods with segmented bodies, numerous walking legs, jointed appendages, and a chitinous exoskeleton. Millipedes have rounded bodies with two pairs of legs per body segment and are mostly slow-moving detritivores that feed on decaying plant material, while centipedes have flatter bodies with one pair of legs per body segment and are fast-moving predators that feed on insects and other small invertebrates.
Subphylum Hexapoda
Kingdom Animalia: Eumetazoa: Bilateria: Nephrozoa: Protostomia: Ecdysozoa: Phylum Arthropoda: Subphylum Hexapoda
Insects are arthropods in Class Insecta, which belongs to Subphylum Hexapoda. Hexapods are defined by having six legs, or three pairs of legs. Insects are by far the largest and most diverse group within Hexapoda, but the subphylum also includes several smaller groups of non-insect hexapods, including springtails (Class Collembola), proturans (Class Protura), and diplurans (Class Diplura). These non-insect hexapods have their mouthparts are enclosed within the head, unlike insects, whose mouthparts are exposed.
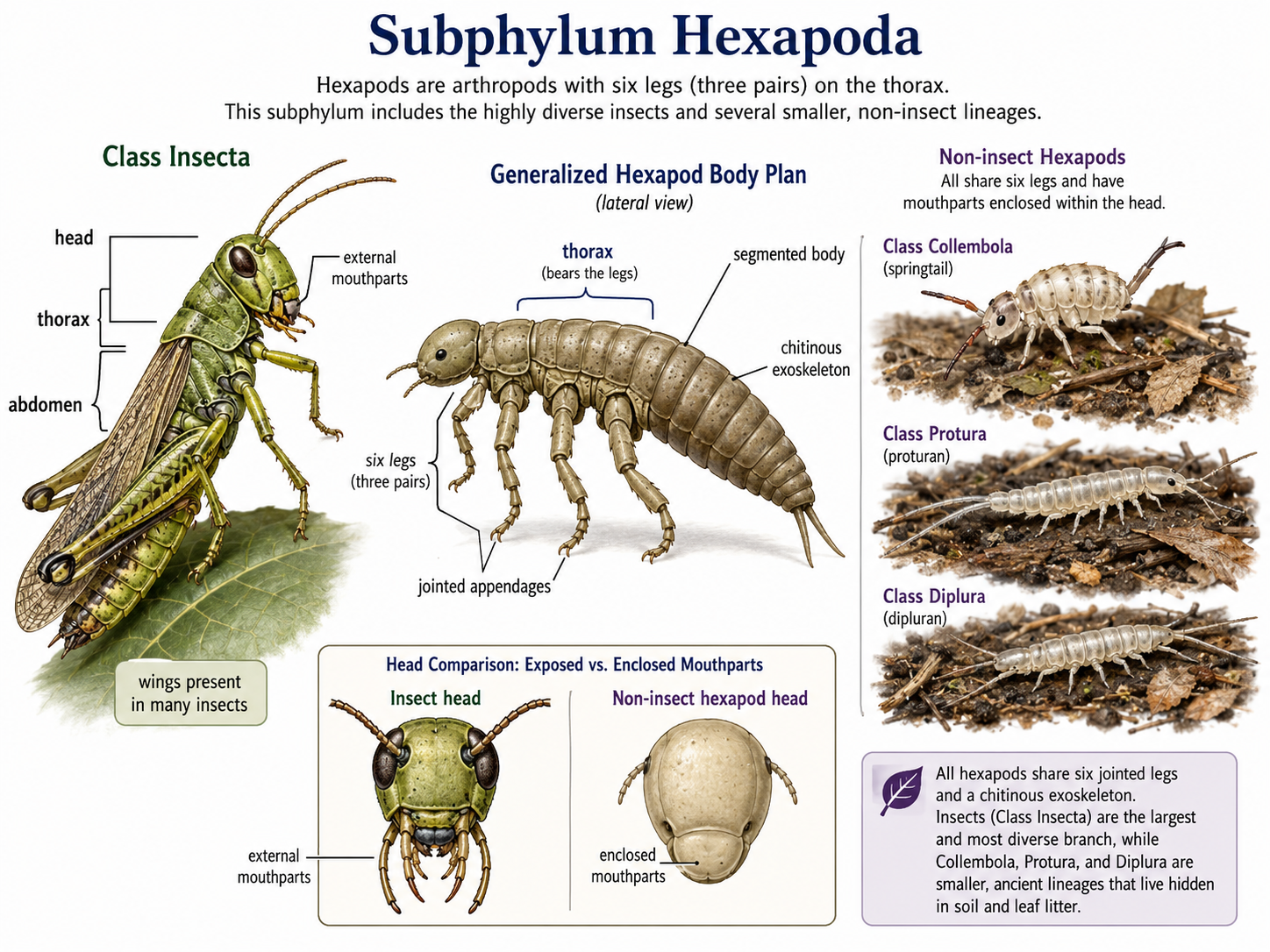
Figure 26. Subphylum Hexapoda. All hexapods share a six-legged body plan, with three pairs of jointed legs attached to the thorax, a segmented body, and a chitinous exoskeleton. This illustration contrasts insects (Class Insecta), which have exposed external mouthparts and often wings, with non-insect hexapods such as springtails (Class Collembola), proturans (Class Protura), and diplurans (Class Diplura), which are small soil- and leaf-litter-dwelling forms with mouthparts enclosed within the head.
Class Insecta (Insects)
Kingdom Animalia: Eumetazoa: Bilateria: Nephrozoa: Protostomia: Ecdysozoa: Phylum Arthropoda: Subphylum Hexapoda: Class Insecta
Insects are arthropods with a chitinous exoskeleton, a body divided into three main regions—the head, thorax, and abdomen—and three pairs of jointed legs attached to the thorax. Most insects also have compound eyes and one pair of antennae, which help them sense light, movement, chemicals, and vibrations in their environment. Insects are the most diverse group of animals on Earth, with more than one million described species and many more still undiscovered. They occupy nearly every terrestrial and freshwater habitat, including forests, grasslands, deserts, wetlands, soils, caves, and human-built environments. Their success comes from several key adaptations, including a protective exoskeleton, small body size, specialized mouthparts, rapid reproduction, metamorphosis, and, in many groups, the ability to fly. Flight allowed insects to escape predators, find food and mates, and colonize new habitats more efficiently than most other invertebrates. Although insects dominate many land ecosystems as pollinators, decomposers, predators, parasites, and prey, relatively few species live in the open ocean, where crustaceans are the dominant arthropods.
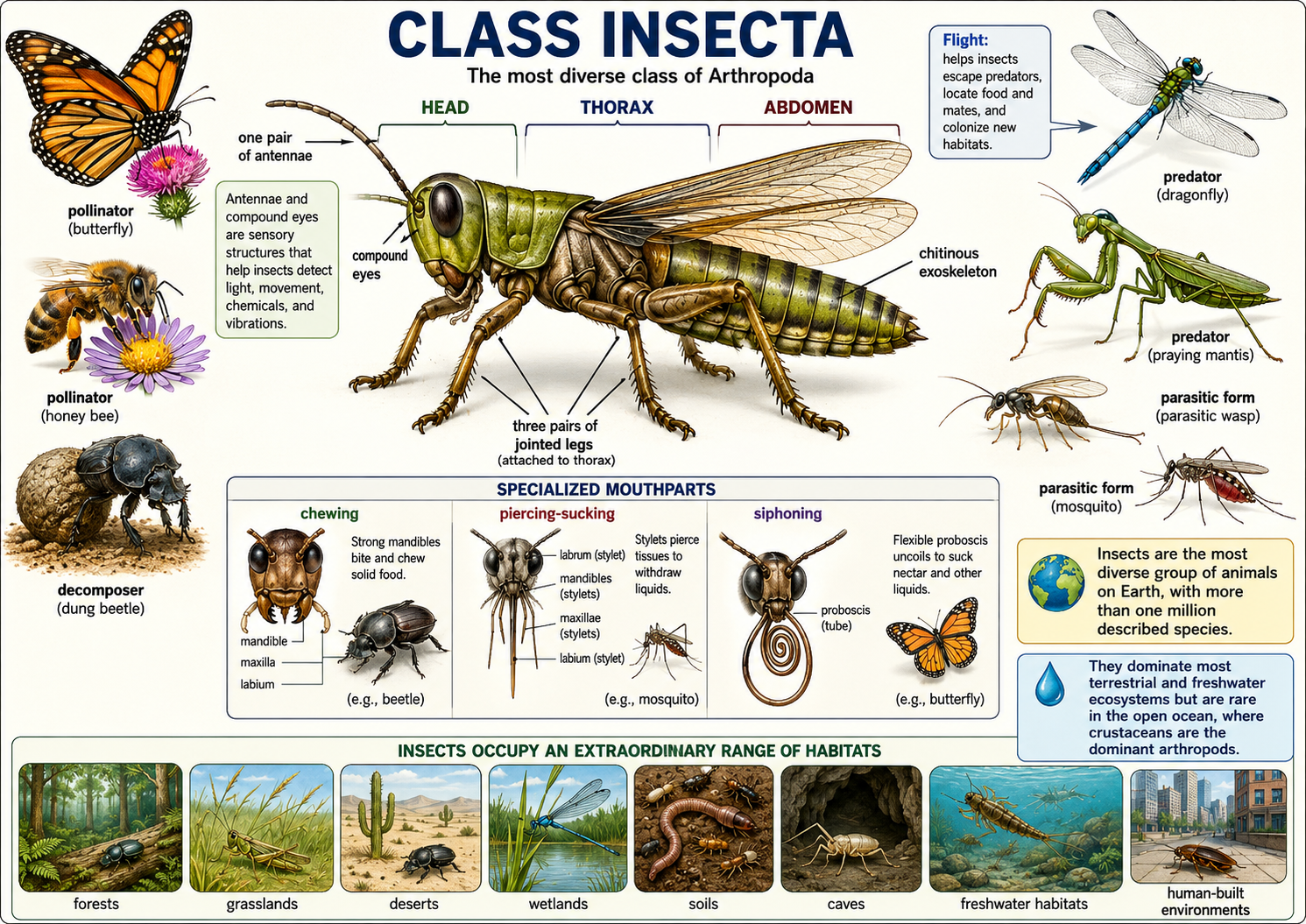
Figure 27. Class Insecta. Insects are arthropods with a head, thorax, abdomen, three pairs of jointed legs, one pair of antennae, compound eyes, specialized mouthparts, and a chitinous exoskeleton. Their diversity and success reflect adaptations such as flight, sensory specialization, varied feeding structures, and roles as pollinators, decomposers, predators, and parasites across most terrestrial and freshwater ecosystems.
Subphylum Chelicerata (Spiders, Scorpions, Ticks, Mites, Horseshoe Crabs)
Kingdom Animalia: Eumetazoa: Bilateria: Nephrozoa: Protostomia: Ecdysozoa: Phylum Arthropoda: Subphylum Chelicerata
Chelicerates (Subphylum Chelicerata) are arthropods that include spiders, scorpions, ticks, mites, horseshoe crabs, and related groups. Their body plan is usually divided into two main regions, or tagmata: the cephalothorax and the abdomen. In some groups, especially mites and ticks, this division is greatly reduced or difficult to see. Chelicerates are named for their chelicerae, the first pair of appendages located near the mouth. These structures are used in feeding and vary widely among groups. In many chelicerates, they function as small pincers or grasping structures. In spiders, the chelicerae are modified into fangs, which are often connected to venom glands and used to subdue prey. Chelicerates also have a second pair of appendages called pedipalps, which may be used for sensing, feeding, grasping, or reproduction. Unlike insects and crustaceans, chelicerates lack antennae and mandibles. Most terrestrial chelicerates, such as spiders and scorpions, have four pairs of walking legs as adults. Marine forms, such as horseshoe crabs, use book gills for gas exchange, while terrestrial forms may use book lungs, tracheae, or both. Their nervous systems are relatively centralized, with many ganglia fused into larger nerve centers in the cephalothorax. Chelicerates rely heavily on sensory hairs and bristles to detect touch, vibrations, air currents, and chemical signals. Many spiders are especially sensitive to vibrations traveling through webs or surfaces, while active hunting spiders often have well-developed eyesight. This combination of specialized appendages, sensory structures, venom in some groups, and efficient predatory behavior has made chelicerates highly successful in both aquatic and terrestrial environments.
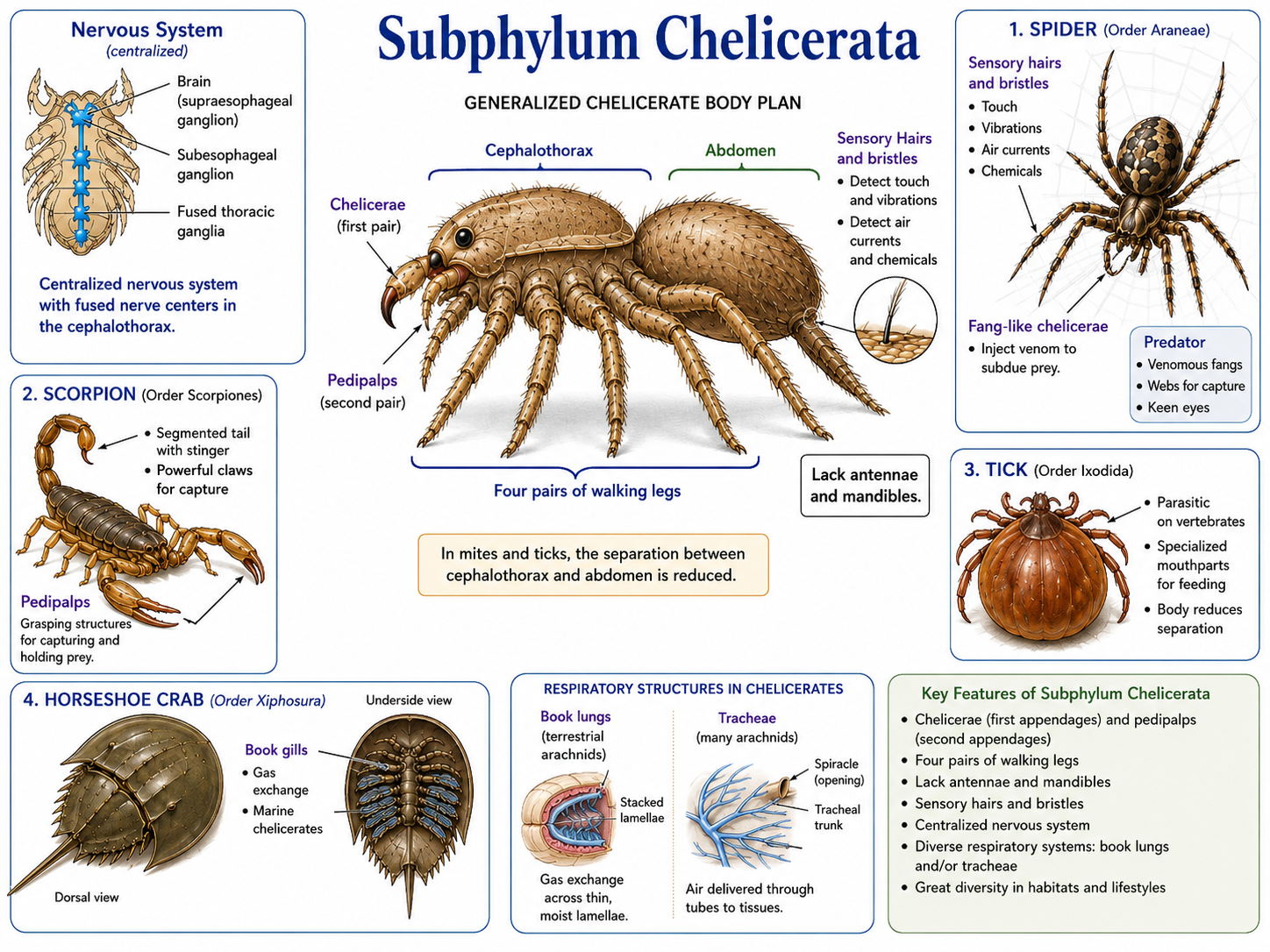
Figure 28. Subphylum Chelicerata. Chelicerates are arthropods characterized by a body typically divided into a cephalothorax and abdomen, four pairs of walking legs in most terrestrial adults, and specialized appendages called chelicerae and pedipalps. This group includes spiders, scorpions, mites, ticks, and horseshoe crabs, and its members are adapted for feeding, sensing, respiration, and predation in both aquatic and terrestrial habitats.
Subphylum Crustacea (Crabs, Lobsters, Shrimp, Barnacles)
Kingdom Animalia: Eumetazoa: Bilateria: Nephrozoa: Protostomia: Ecdysozoa: Phylum Arthropoda: Subphylum Crustacea
Crustaceans (Subphylum Crustacea) are a diverse group of arthropods that include crabs, lobsters, shrimp, crayfish, barnacles, krill, copepods, and many other forms. They are primarily aquatic, especially marine, although some live in freshwater and a few groups, such as pill bugs (better known as rollie pollies), have adapted to life on land. Crustaceans are especially important in aquatic ecosystems, where many species function as predators, scavengers, filter feeders, grazers, and plankton. Like other arthropods, crustaceans have a segmented body, jointed appendages, and a hard exoskeleton made of chitin. In many larger crustaceans, such as crabs and lobsters, the exoskeleton is strengthened with calcium carbonate, making it thick and protective. Because the exoskeleton does not grow with the animal, crustaceans must periodically shed it through molting. The crustacean body is often divided into two tagmata: the cephalothorax and the abdomen. The cephalothorax forms from the fusion of the head and thorax and is often covered by a broad protective shield called the carapace. This carapace protects many of the internal organs and may also cover the gills in aquatic species. Unlike chelicerates, crustaceans have antennae. In fact, they typically have two pairs of antennae, which are used for sensing chemicals, touch, and movement in the environment. Crustaceans also possess mandibles, jaw-like mouthparts used for biting, crushing, or processing food. Their appendages are highly modified for different jobs, including walking, swimming, feeding, sensing, defense, and reproduction. Most crustaceans breathe using gills, which allow them to exchange gases in water. Many also have compound eyes, often mounted on movable stalks, giving them a wide field of view. This combination of a protective exoskeleton, specialized appendages, sensory antennae, and aquatic adaptations has made crustaceans one of the most successful arthropod groups in marine and freshwater environments.
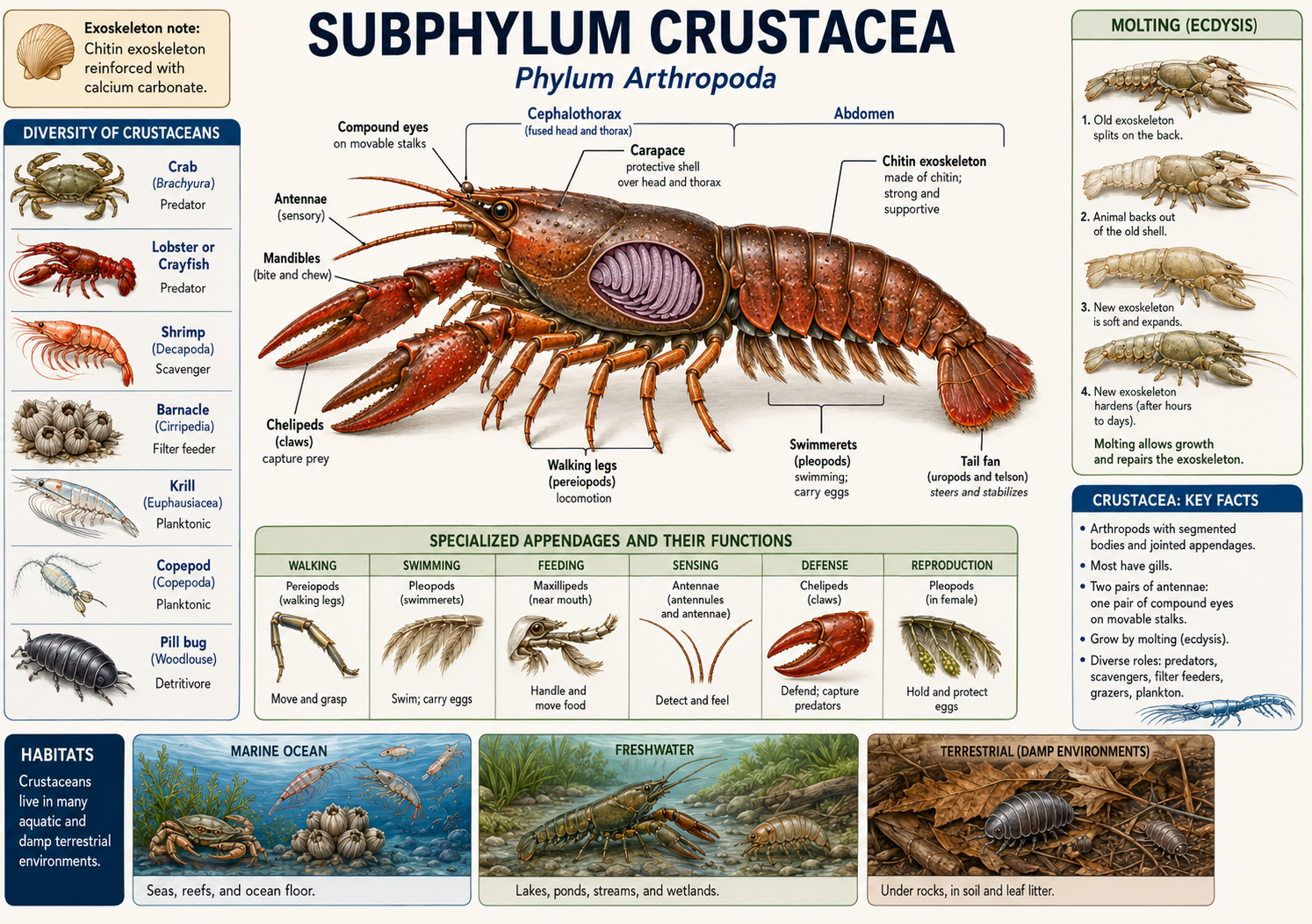
Figure 29. Subphylum Crustacea. Crustaceans are arthropods with a segmented body, jointed appendages, a chitinous exoskeleton, two pairs of antennae, and mandibles for feeding. Most are aquatic and breathe with gills, and their appendages and body regions are highly specialized for diverse roles in marine, freshwater, and, in a few cases, terrestrial environments.
Deuterostomes
Kingdom Animalia: Eumetazoa: Bilateria: Nephrozoa: Deuterostomia
Deuterostomes (Clade Deuterostomia) are one of the two major branches of Nephrozoa, the clade of bilaterian animals with a through-gut and complex organ systems. Compared with protostomes, deuterostomes contain fewer species, but they include some of the most familiar and biologically important animals on Earth, including sea stars, sea urchins, acorn worms, tunicates, fishes, amphibians, reptiles, birds, and mammals. The defining developmental feature of deuterostomes occurs early in embryonic development: the blastopore becomes the anus, while the mouth forms second. This contrasts with protostomes, in which the blastopore typically becomes the mouth. Deuterostomes are divided into two major evolutionary lineages: Ambulacraria and Chordata. Although these groups may appear very different as adults, they share deep developmental and evolutionary ancestry.
Ambulacraria
Kingdom Animalia: Eumetazoa: Bilateria: Nephrozoa: Deuterostomia: Ambulacraria
Ambulacrarians (Clade Ambulacraria) include Phylum Echinodermata and Phylum Hemichordata. Echinoderms include sea stars, brittle stars, sea urchins, sand dollars, sea cucumbers, and crinoids. Although adult echinoderms often show five-part radial symmetry, their larvae are bilaterally symmetrical, revealing their placement within Bilateria. Echinoderms also possess a unique water vascular system, which helps power tube feet used in movement, feeding, and gas exchange. Hemichordates, including acorn worms and pterobranchs, are mostly marine animals that retain a more obviously worm-like body plan. Together, echinoderms and hemichordates show that deuterostomes include more than vertebrates; they also include marine invertebrate lineages with distinctive body plans and ecological roles.
Chordata
Kingdom Animalia: Eumetazoa: Bilateria: Nephrozoa: Deuterostomia: Chordata
Chordates (Phylum Chordata) include tunicates, lancelets, and vertebrates. All chordates share several key features at some point in life: a notochord, a dorsal hollow nerve cord, pharyngeal slits or pouches, and a post-anal tail. In some chordates, such as lancelets, these traits remain visible throughout life. In others, such as tunicates, they are most obvious in the larval stage. Vertebrates are the most familiar chordates and include fishes, amphibians, reptiles, birds, and mammals. In vertebrates, the notochord is largely replaced by the vertebral column, and the nervous system becomes highly centralized. Chordates demonstrate how the deuterostome body plan gave rise to active swimmers, complex nervous systems, internal skeletons, and eventually the terrestrial vertebrate lineages.
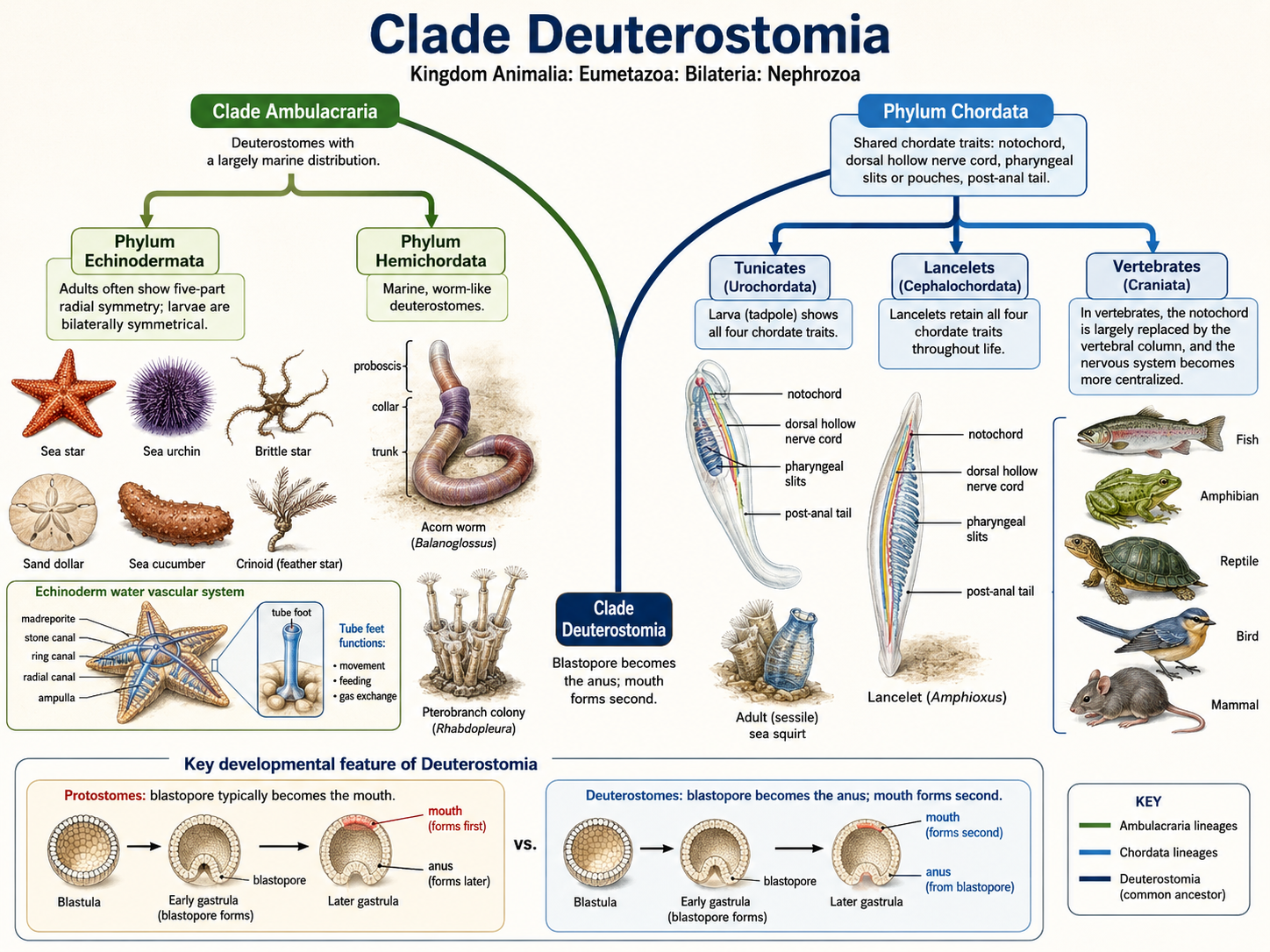
Figure 30. Deuterostomia. Deuterostomes are bilaterian animals within Nephrozoa whose embryonic blastopore becomes the anus while the mouth forms second. The clade includes two major lineages: Ambulacraria, containing Phylum Echinodermata and Phylum Hemichordata, and Phylum Chordata, containing tunicates, lancelets, and vertebrates.
Echinodermata
Kingdom Animalia: Eumetazoa: Bilateria: Nephrozoa: Deuterostomia: Ambulacraria: Phylum Echinodermata
Echinoderms are marine animals that include sea stars, brittle stars, sea urchins, sand dollars, sea cucumbers, and crinoids. The name Echinodermata means “spiny skin,” referring to the hard plates, spines, or bumps found in many members of the group. Although echinoderms may look very different from chordates, they belong to Clade Deuterostomia, the same major evolutionary branch that includes vertebrates.
Echinodermata: Bilateral Larvae, Radial Adult
Kingdom Animalia: Eumetazoa: Bilateria: Nephrozoa: Deuterostomia: Ambulacraria: Phylum Echinodermata
One of the strangest features of echinoderms is their symmetry. Adult echinoderms often show radial symmetry, usually arranged around five body sections. Sea stars, for example, commonly have five arms around a central body. However, echinoderms are not closely related to radially symmetrical animals such as cnidarians. They are members of Clade Bilateria, the group of animals with bilaterally symmetrical ancestry. This is clearest in their larvae, which are bilaterally symmetrical before transforming into radially symmetrical adults.
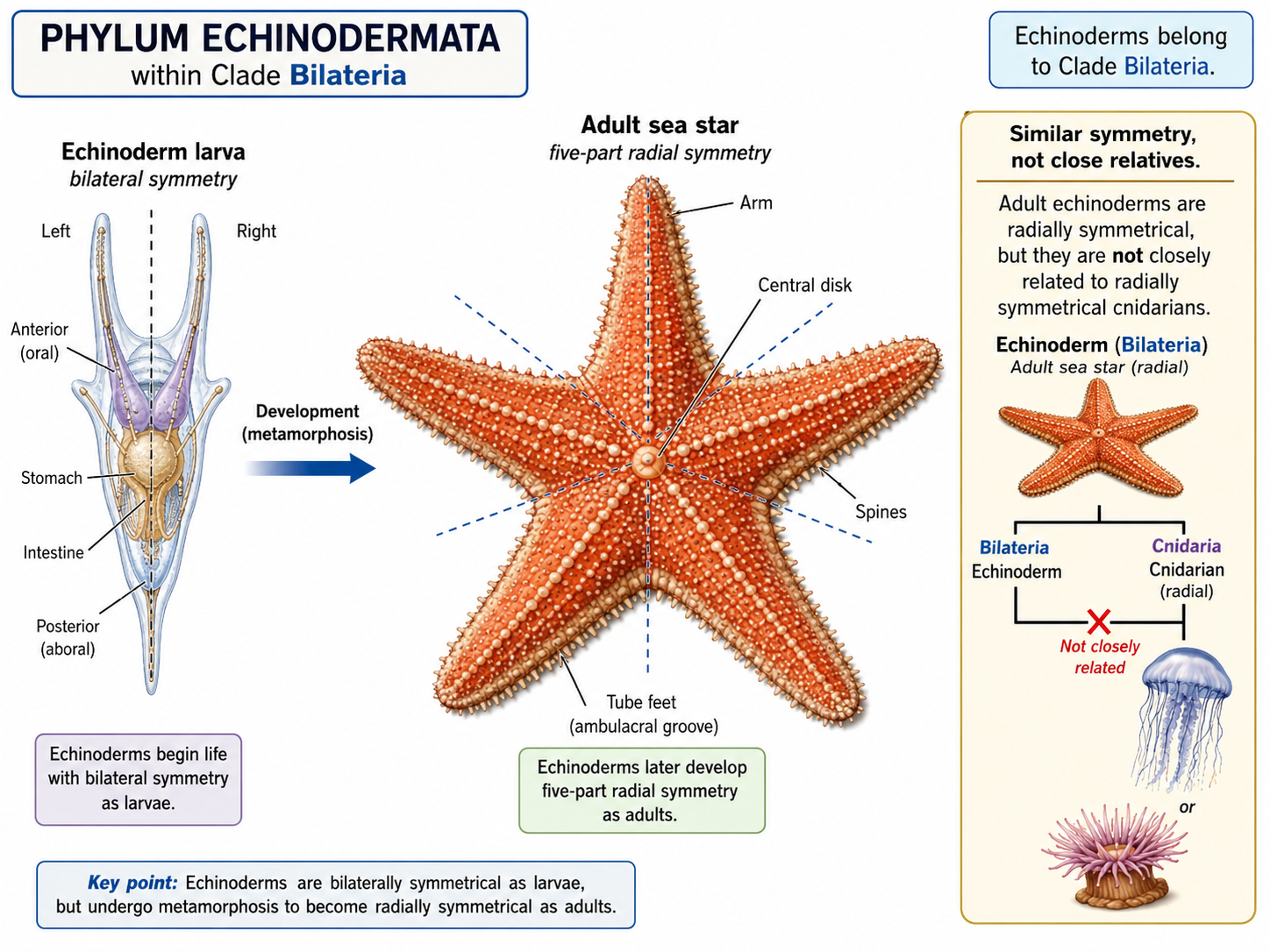
Figure 31. Echinoderm symmetry. Adult echinoderms, such as sea stars, often show five-part radial symmetry, but they belong to Clade Bilateria because their larvae are bilaterally symmetrical. This developmental pattern shows that echinoderm radial symmetry evolved secondarily from bilaterian ancestry.
Echinodermata: Endoskeleton
Kingdom Animalia: Eumetazoa: Bilateria: Nephrozoa: Deuterostomia: Ambulacraria: Phylum Echinodermata
Echinoderms also have an internal skeleton called an endoskeleton. This skeleton is made of calcium carbonate plates located beneath the skin. In sea urchins and sand dollars, these plates are fused into a rigid shell-like structure. In sea stars and sea cucumbers, the plates are smaller or more flexible. The endoskeleton provides support and protection, and in many species it bears spines that help defend the animal from predators.
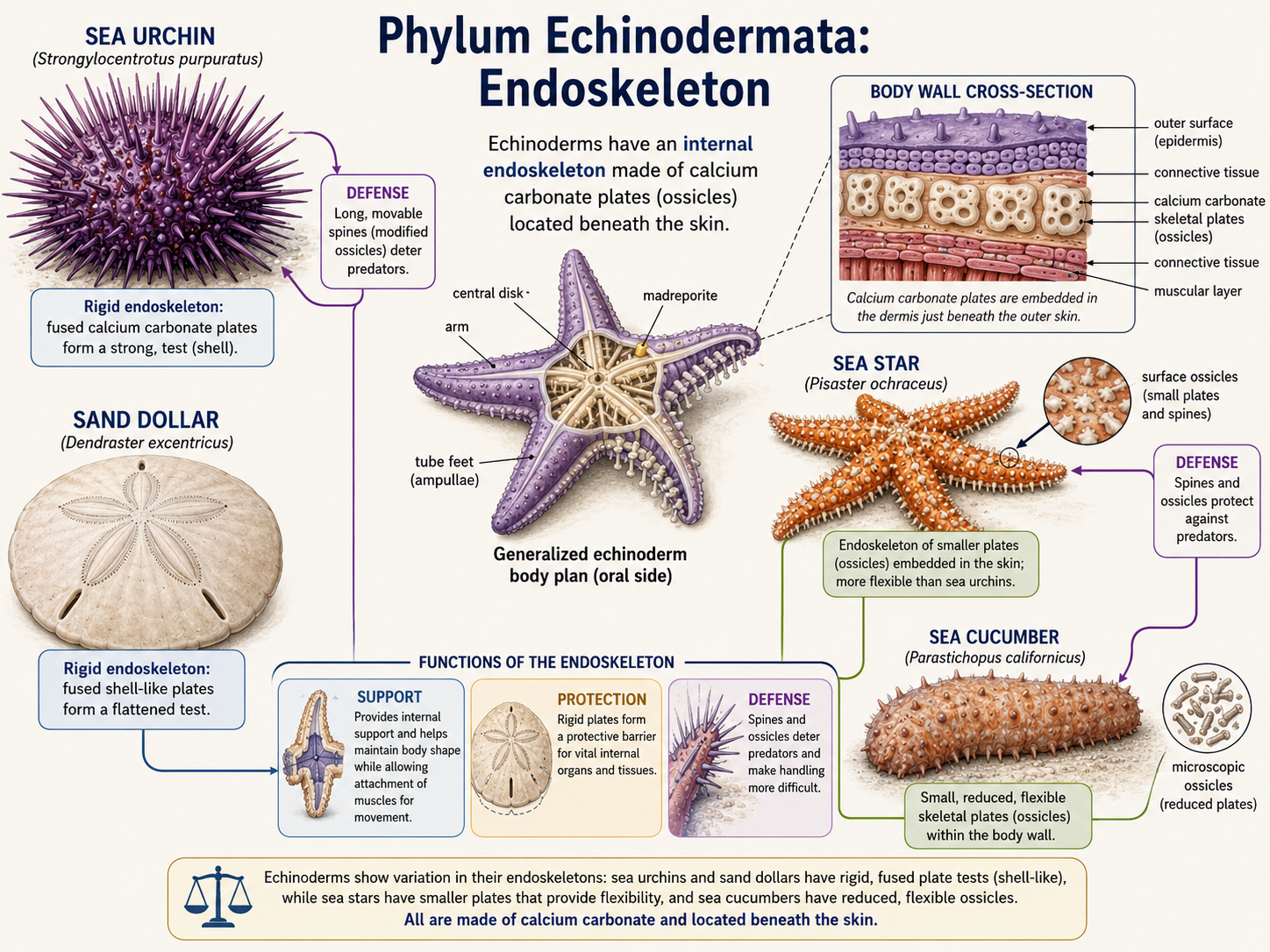
Figure 32. Echinoderm endoskeleton. Echinoderms possess an internal skeleton made of calcium carbonate plates located beneath the skin. In some groups, such as sea urchins and sand dollars, the plates are fused into a rigid structure, while in others, such as sea stars and sea cucumbers, they are smaller or more flexible; together, these plates provide support, protection, and often bear defensive spines.
Echinodermata: Water Vascular System
Kingdom Animalia: Eumetazoa: Bilateria: Nephrozoa: Deuterostomia: Ambulacraria: Phylum Echinodermata
The most distinctive feature of echinoderms is the water vascular system, a network of fluid-filled canals used in movement, feeding, and gas exchange. In sea stars, this system connects to many small extensions called tube feet. By changing fluid pressure inside the system, the animal can extend and retract its tube feet. Tube feet allow sea stars to move, grip prey, attach to surfaces, and handle food.
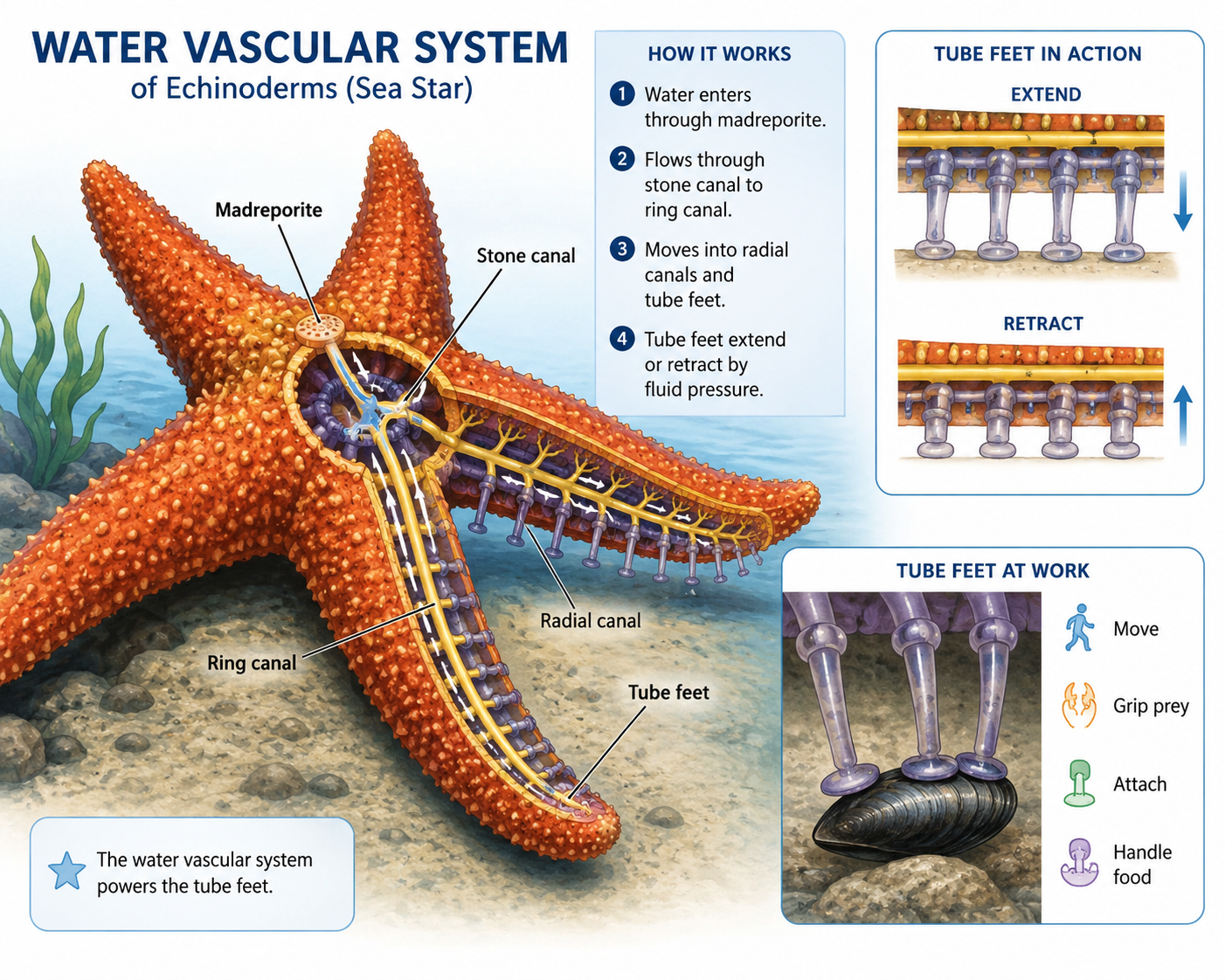
Figure 33. Water vascular system in echinoderms. Echinoderms possess a unique water vascular system, a network of fluid-filled canals that helps power tube feet. In sea stars, the tube feet are used for movement, feeding, gas exchange, attachment, and gripping prey by changing fluid pressure within the system.
Major Echinodermata Lineages
Kingdom Animalia: Eumetazoa: Bilateria: Nephrozoa: Deuterostomia: Ambulacraria: Phylum Echinodermata
Phylum Echinodermata includes five major living classes: Class Crinoidea, Class Asteroidea, Class Ophiuroidea, Class Echinoidea, and Class Holothuroidea.
Class Crinoidea
Kingdom Animalia: Eumetazoa: Bilateria: Nephrozoa: Deuterostomia: Ambulacraria: Phylum Echinodermata: Class Crinoidea
Class Crinoidea includes the feather stars and sea lilies, some of the most ancient-looking living echinoderms. Unlike many other echinoderms, crinoids usually hold their mouth facing upward, toward the water column, rather than downward toward the seafloor. They are mostly suspension feeders, meaning they capture tiny food particles, plankton, and organic material drifting in the water. Crinoids feed with long, branching, feathery arms covered with many small side branches called pinnules. These arms form a filtering surface that catches suspended food particles. Cilia and mucus help move captured food along grooves in the arms toward the mouth. Sea lilies are usually attached to the seafloor by a stalk, giving them a plant-like appearance, although they are animals. Feather stars lack a long adult stalk and are more mobile. They can cling to surfaces with small grasping structures, crawl slowly, and in some cases swim short distances by waving their arms. Crinoids are especially common in marine habitats with steady water movement, where currents bring food particles within reach. Their body form reflects a feeding strategy built around staying in position and filtering the surrounding water.
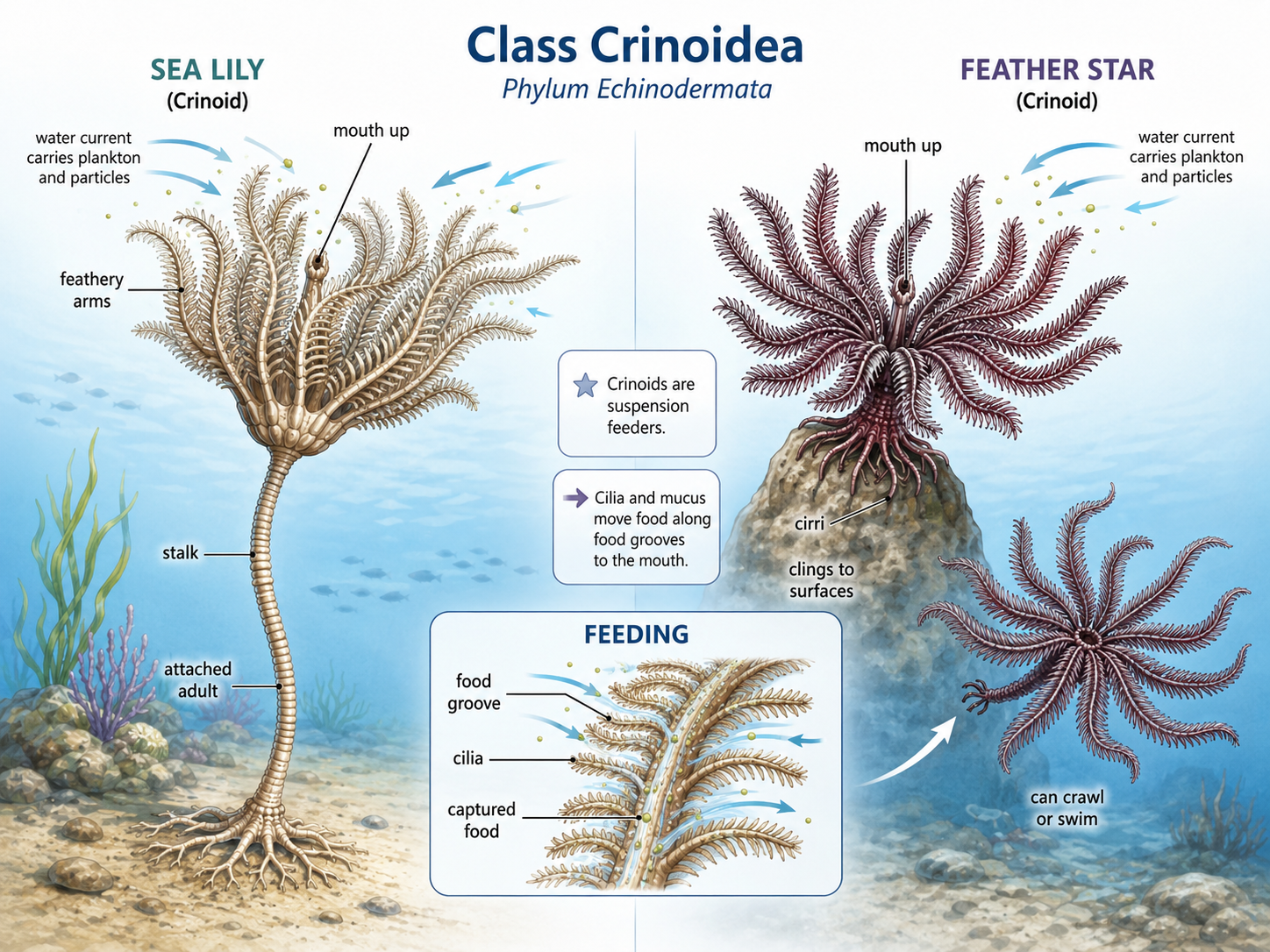
Figure 34. Class Crinoidea. Crinoids, including sea lilies and feather stars, are echinoderms adapted for suspension feeding. Their upward-facing mouth and branched, feathery arms with pinnules allow them to capture food particles from the water, while sea lilies remain attached by a stalk and feather stars are more mobile, using cirri to cling, crawl, or sometimes swim.
Class Asteroidea
Kingdom Animalia: Eumetazoa: Bilateria: Nephrozoa: Deuterostomia: Ambulacraria: Phylum Echinodermata: Class Asteroidea
Class Asteroidea includes the sea stars, which are often called starfish, although they are not fish. Most sea stars have five arms arranged around a central body, but some species have many more. In sea stars, the arms usually merge broadly with the central disc, so the boundary between the arms and body is not as sharply defined as it is in brittle stars. Sea stars are among the best-known predatory echinoderms. Many species live in rocky intertidal zones and shallow marine habitats, where they feed on mussels, clams, oysters, snails, barnacles, and other invertebrates. Their tube feet, powered by the water vascular system, allow them to move slowly but with great force. When feeding on bivalves such as mussels or clams, some sea stars use their tube feet to pull the shell open slightly, then extend part of the stomach outside the body to digest the prey externally. Sea stars also have strong regenerative abilities. Many can regrow lost arms, and in some species a detached arm can regenerate much of the body if part of the central disc remains. Ecologically, sea stars can be important predators that influence the structure of marine communities by controlling the abundance of prey species.

Figure 35. Class Asteroidea. Sea stars are predatory echinoderms with broad arms, tube feet powered by the water vascular system, and strong regenerative abilities. Many species feed on bivalves and other invertebrates, sometimes using external digestion, and they often play an important ecological role by influencing the abundance of prey in marine communities.
Class Ophiuroidea
Kingdom Animalia: Eumetazoa: Bilateria: Nephrozoa: Deuterostomia: Ambulacraria: Phylum Echinodermata: Class Asteroidea
Class Ophiuroidea includes brittle stars and basket stars. These echinoderms have a small, distinct central disc and long, thin arms that are clearly set off from the body. This gives them a very different appearance from sea stars, even though both groups are echinoderms. Brittle stars are usually very flexible and move quickly compared with many other echinoderms. Instead of relying mainly on tube feet for movement, they move by rowing, bending, or snakelike motions of their arms. Their tube feet are often used more for feeding and sensing than for powerful locomotion. Many brittle stars live under rocks, in crevices, on coral reefs, or buried in sediment, where their slender arms can reach out into the surrounding environment. Ophiuroids show a wide range of feeding strategies. Some brittle stars are scavengers that consume dead organic material, some are deposit feeders that collect food from sediment, and others are suspension feeders that capture particles from the water. Basket stars are specialized suspension feeders with highly branched arms that form a net-like feeding structure. These branching arms increase surface area and help capture plankton and suspended organic particles from passing currents.

Figure 36. Class Ophiuroidea. Ophiuroids, including brittle stars and basket stars, are echinoderms with a small central disc and long, slender arms that are sharply distinct from the body. Brittle stars move mainly by flexible arm movements and use tube feet largely for sensing and feeding, while basket stars use their highly branched arms as a net-like surface for suspension feeding.
Class Echinoidea (Urchins and Sand Dollars.
Kingdom Animalia: Eumetazoa: Bilateria: Nephrozoa: Deuterostomia: Ambulacraria: Phylum Echinodermata: Class Echinoidea
Class Echinoidea includes sea urchins, heart urchins, and sand dollars. Unlike sea stars and brittle stars, echinoids do not have obvious arms. Their body is enclosed in a rigid internal skeleton called a test, made of fused calcium carbonate plates. This test gives many echinoids a firm, compact body shape. Sea urchins usually have rounded bodies covered with movable spines. They move using both their spines and tube feet. Many sea urchins are herbivores that graze on algae, kelp, and other plant-like marine growth. In kelp forest ecosystems, sea urchins can play a major ecological role. When their populations are balanced, they help control algal growth. When their populations become too large, they can overgraze kelp forests and create areas known as urchin barrens. Many sea urchins feed using a complex jaw-like structure called Aristotle’s lantern, which allows them to scrape algae from rocks or process other food. Sand dollars are flattened echinoids adapted for life in sandy or muddy sediment. They often live partly buried and feed on small organic particles. Their tube feet, small spines, and cilia help move food toward the mouth. Their flattened body shape helps them remain stable in shifting sediment and shallow-water environments.

Figure 37. Class Echinoidea. Echinoids, including sea urchins, heart urchins, and sand dollars, are echinoderms with a rigid test made of fused calcium carbonate plates and no obvious arms. Sea urchins use spines, tube feet, and Aristotle’s lantern for movement and feeding, while sand dollars are flattened for life in sediment, where they feed on small organic particles and remain stable in shifting substrates.
Class Holothuroidea
Kingdom Animalia: Eumetazoa: Bilateria: Nephrozoa: Deuterostomia: Ambulacraria: Phylum Echinodermata: Class Holothuroidea
Class Holothuroidea includes the sea cucumbers. Sea cucumbers look very different from sea stars, sea urchins, and brittle stars because their bodies are elongated and soft. Their calcium carbonate skeletal plates are greatly reduced and embedded in the body wall, so they lack the rigid, obvious skeleton seen in sea urchins or sand dollars. Most sea cucumbers live on or within the seafloor sediment. Many are deposit feeders, meaning they ingest sediment and extract organic material, bacteria, and tiny food particles from it. In this way, they act somewhat like marine earthworms, helping process sediment and recycle nutrients. Other sea cucumbers are suspension feeders that use branching feeding tentacles to capture particles from the water. The tentacles around the mouth of a sea cucumber are modified tube feet, showing how the echinoderm water vascular system has been adapted for feeding. Some sea cucumbers also have unusual defense mechanisms. Certain species can release sticky internal structures called Cuvierian tubules, while others may expel parts of their internal organs when threatened and later regenerate them. Although sea cucumbers may not look much like other echinoderms, their tube feet, water vascular system, internal skeletal plates, and evolutionary relationships clearly place them within Phylum Echinodermata. Their unusual body form is a specialized adaptation to life on and within marine sediments.
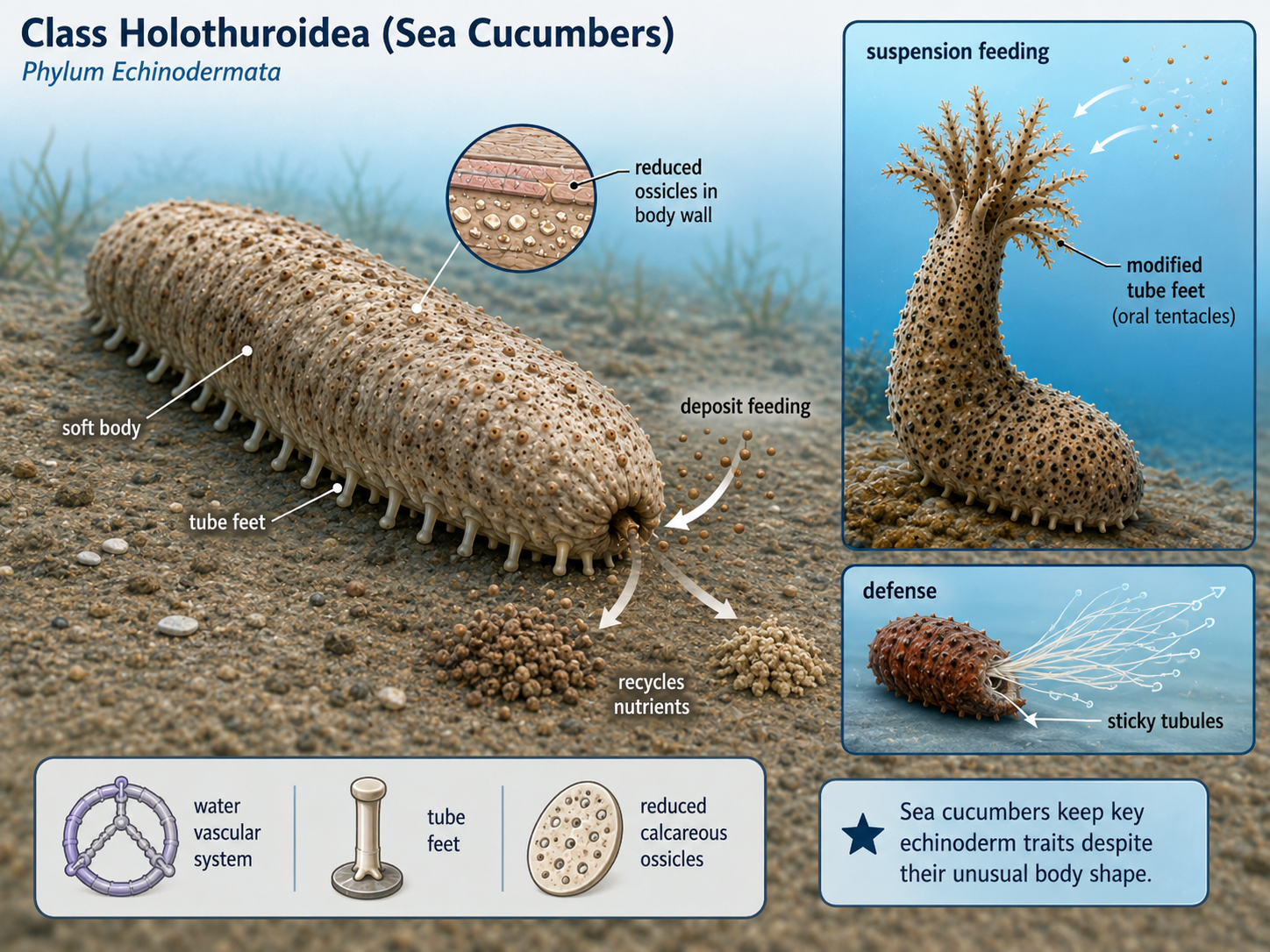
Figure 38. Class Holothuroidea. Sea cucumbers are echinoderms with elongated, soft bodies and greatly reduced skeletal plates embedded in the body wall. Many are deposit feeders that process sediment and recycle nutrients, while others are suspension feeders that use branching tentacles, modified from tube feet, to capture particles from the water; some also possess unusual defenses such as Cuvierian tubules or the ability to expel and later regenerate internal organs.
Phylum Chordata
Kingdom Animalia: Eumetazoa: Bilateria: Nephrozoa: Deuterostomia: Phylum Chordata
Chordates are animals in Phylum Chordata, a major branch of the deuterostomes. This phylum includes vertebrates such as fishes, amphibians, reptiles, birds, and mammals, but not all chordates are vertebrates. Some chordates, including lancelets and tunicates, do not have a backbone as adults. These invertebrate chordates are important because they help reveal the early evolution of the chordate body plan. All chordates share several key traits at some point in their life cycle. These traits may be visible in the adult, or they may appear only during embryonic or larval development. The major chordate features are a notochord, a dorsal hollow nerve cord, pharyngeal slits or pouches, and a post-anal tail. The notochord is a flexible support rod that runs along the body. In lancelets and some other nonvertebrate chordates, it remains the main support structure. In vertebrates, it is mostly replaced by the vertebral column, or backbone, although remnants remain between the vertebrae. This structure was important because it supported the body while still allowing flexible movement. Chordates also have a dorsal hollow nerve cord, which runs along the back of the body and forms as a tube. In vertebrates, this structure develops into the brain and spinal cord. This differs from many protostomes, such as annelids and arthropods, which usually have solid nerve cords on the underside of the body. Another chordate feature is the presence of pharyngeal slits or pouches, which form in the throat region. In many aquatic chordates, these structures are used in feeding or gas exchange. In fishes, they develop into gills. In terrestrial vertebrates, including humans, they do not become functional gills. Instead, related embryonic structures contribute to parts of the jaw, ear, throat, and neck. Chordates also have a post-anal tail, meaning a tail that extends beyond the anus. In many aquatic chordates, this tail helps with swimming. In humans and other apes, the tail appears during embryonic development but is reduced before birth, leaving the coccyx, or tailbone.
Together, these features define Phylum Chordata. However, chordates have diversified into very different body forms. The three major living chordate lineages are Subphylum Cephalochordata, Subphylum Tunicata, and Subphylum Vertebrata.
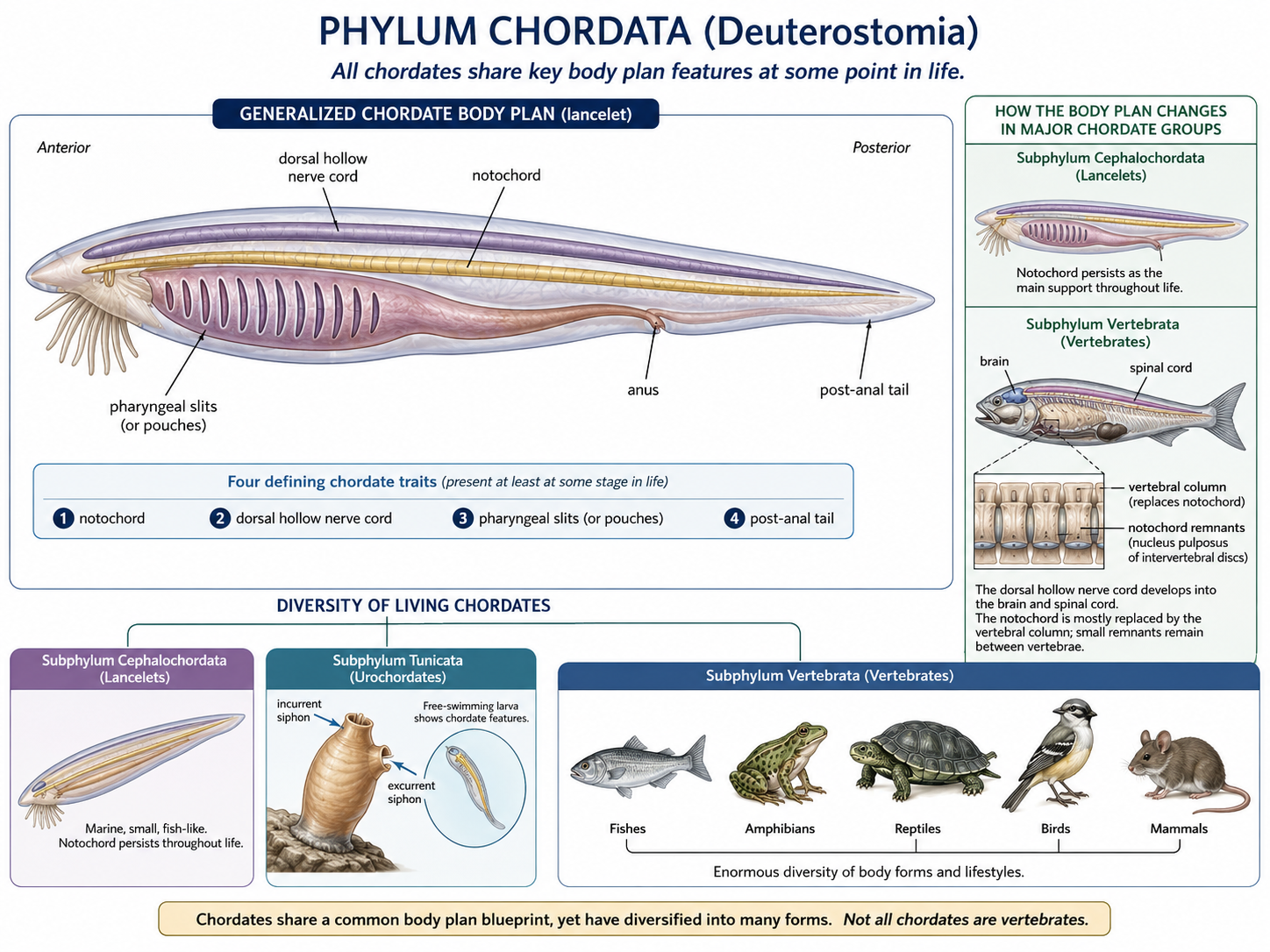
Figure 39. Phylum Chordata. All chordates share four key traits at some stage in life: a notochord, dorsal hollow nerve cord, pharyngeal slits or pouches, and post-anal tail. Lancelets, tunicates, and vertebrates all reflect this shared body plan, although in vertebrates the notochord is mostly replaced by the vertebral column and the nerve cord develops into the brain and spinal cord.
Subphylum Cephalochordata: Lancelets
Kingdom Animalia: Eumetazoa: Bilateria: Nephrozoa: Deuterostomia: Phylum Chordata: Subphylum Cephalochordata
Subphylum Cephalochordata includes the lancelets. Lancelets are small, fish-shaped marine animals that usually live partly buried in sand, with only the front end of the body exposed. They are suspension feeders, drawing water into the mouth and filtering tiny food particles from it. Lancelets are not fishes, and they do not have a skull, jaws, vertebrae, or paired fins. However, they show the basic chordate body plan very clearly. As adults, they retain a notochord, dorsal hollow nerve cord, pharyngeal slits, and post-anal tail. Because these traits remain visible throughout life, lancelets are useful models for understanding what early chordates may have been like. The lancelet notochord extends through most of the body and provides support during swimming. Their muscles are arranged in repeated blocks, allowing them to move side to side in a fishlike motion. Although lancelets are simple compared with vertebrates, they are not “primitive” in the sense of being unchanged or inferior. They are living animals with their own evolutionary history. Still, their anatomy preserves many features that help scientists study the origin of the chordate body plan.
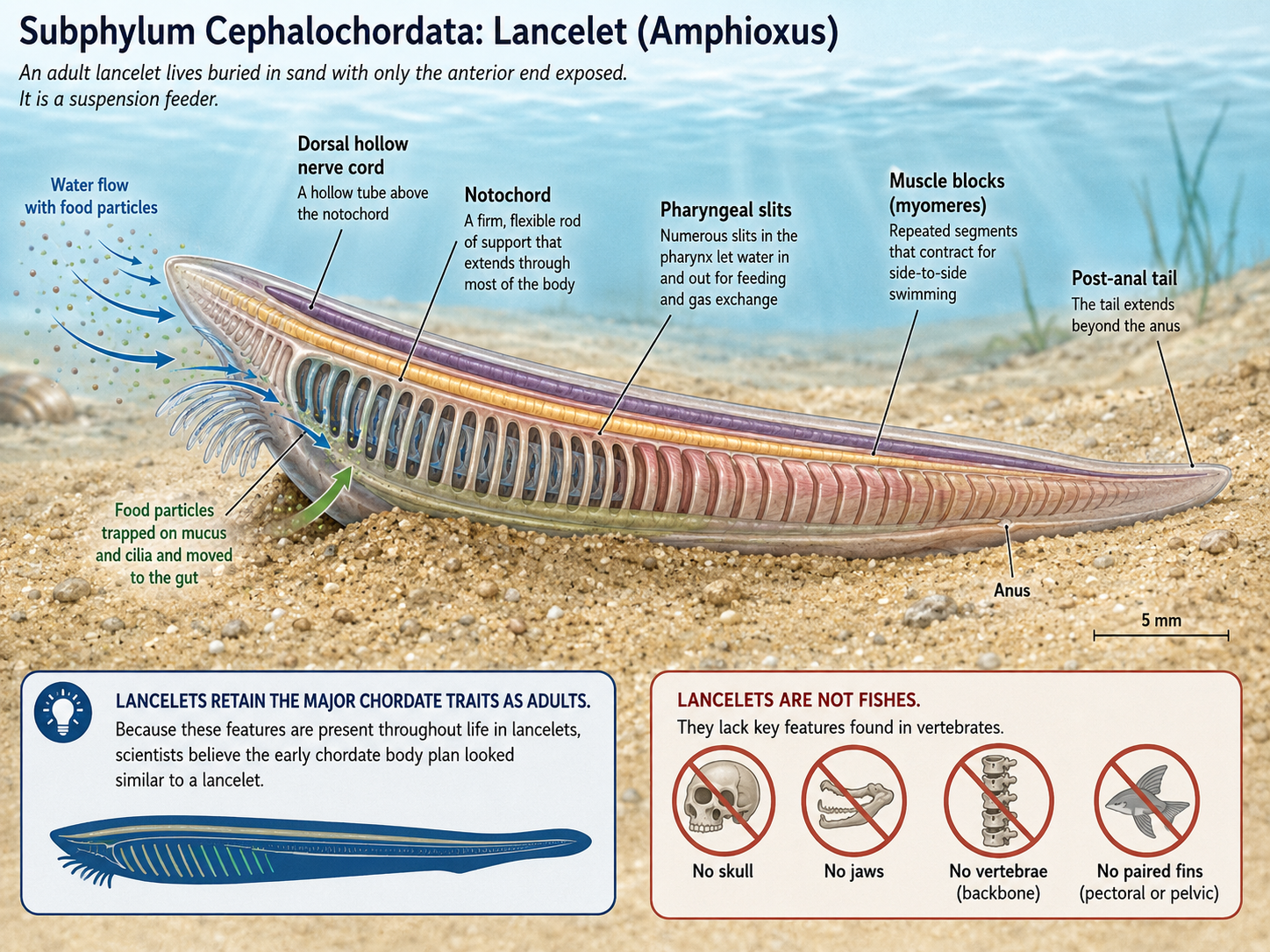
Figure 40. Subphylum Cephalochordata. A lancelet lives partly buried in marine sand and suspension feeds by drawing water and food particles into the pharynx. Adult lancelets retain major chordate traits, including a notochord, dorsal hollow nerve cord, pharyngeal slits, muscle blocks, and post-anal tail. Although lancelets resemble small fishes, they are not vertebrates and lack a skull, jaws, vertebrae, and paired fins.
Subphylum Tunicata: Tunicates or Sea Squirts
Kingdom Animalia: Eumetazoa: Bilateria: Nephrozoa: Deuterostomia: Phylum Chordata: Subphylum Tunicata
Subphylum Tunicata, also called Urochordata, includes the tunicates. Many tunicates are commonly called sea squirts. These marine animals often look nothing like typical chordates as adults, but their larvae reveal their chordate identity. Most adult tunicates are sessile suspension feeders, meaning they attach to a surface and filter food particles from seawater. Water enters through an incurrent siphon, passes through a large pharyngeal basket where food particles are trapped, and exits through an excurrent siphon. This gives many tunicates a body plan with two visible openings. The adult body is surrounded by a tough outer covering called a tunic, which contains a cellulose-like material called tunicin. This makes tunicates unusual among animals, because cellulose-like structural materials are more commonly associated with plants and algae. Tunicate larvae are much more obviously chordate-like than the adults. A typical tunicate larva has a notochord, dorsal hollow nerve cord, and post-anal tail, which help it swim briefly before settling onto a surface. After attachment, many tunicates undergo a dramatic metamorphosis. The tail and notochord are reduced, and the adult becomes specialized for suspension feeding. This life cycle shows an important point about chordates: the defining chordate traits do not always remain visible in the adult body. Tunicates are also important evolutionarily because molecular evidence shows that they are closely related to vertebrates. Although adult tunicates may look simple, they are not outside the chordate story. They are one of the closest living relatives of vertebrates and provide important clues about how vertebrate traits evolved.
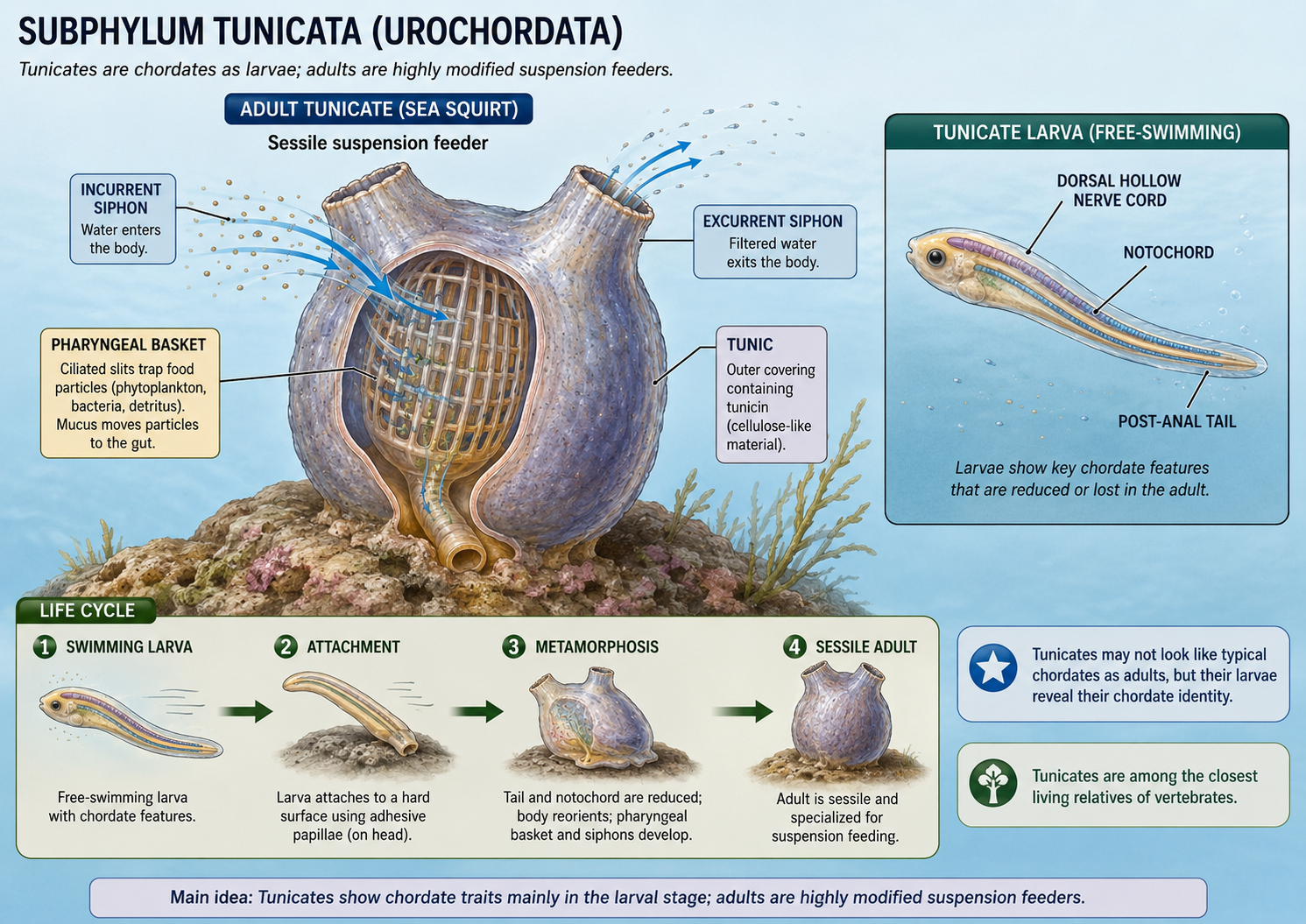
Figure 41. Tunicate body plan and larval chordate traits. Adult tunicates are sessile suspension feeders that draw water through an incurrent siphon, filter food in the pharyngeal basket, and release water through an excurrent siphon. Although adults are highly modified, the swimming larva shows chordate traits, including a notochord, dorsal hollow nerve cord, and post-anal tail. During metamorphosis, the tail and notochord are reduced as the animal becomes a sessile adult.
Subphylum Vertebrata: Vertebrates
Kingdom Animalia: Eumetazoa: Bilateria: Nephrozoa: Deuterostomia: Phylum Chordata: Subphylum Vertebrata
Subphylum Vertebrata includes the chordates with a vertebral column, or backbone. Vertebrates include fishes, amphibians, reptiles, birds, and mammals. Vertebrates are chordates, but they have additional traits that distinguish them from lancelets and tunicates. One of the most important vertebrate traits is the development of a cranium, or skull, which protects the brain and major sensory organs. Vertebrates also have a more complex brain, paired sense organs, a closed circulatory system with a muscular heart, and an internal skeleton. In most vertebrates, the notochord is replaced or surrounded by vertebrae, which protect the spinal cord and provide strong internal support. The vertebrate body plan allowed animals to become larger, more active, and more behaviorally complex. Over evolutionary time, vertebrates evolved several major innovations, including mineralized tissues, jaws, paired appendages, bony skeletons, lungs or lung-like organs, limbs, amniotic eggs, feathers, hair, and placentas. These innovations did not appear all at once. They accumulated over hundreds of millions of years, producing the enormous diversity of vertebrates alive today.

Figure 42. Subphylum Vertebrata. Vertebrate body plan and evolutionary innovations. Vertebrates are chordates distinguished by a vertebral column, cranium, internal skeleton, muscular heart, and closed circulatory system. The backbone supports the body and protects the spinal cord, while the skull protects the brain and paired sense organs. Over time, innovations such as mineralized tissues, jaws, paired appendages, lungs, limbs, amniotic eggs, feathers, hair, and placentas evolved step by step, producing the diversity of modern fishes, amphibians, reptiles, birds, and mammals.
Superclass Cyclostomi: Jawless Vertebrates (Hagfish and Lampreys)
Kingdom Animalia: Eumetazoa: Bilateria: Nephrozoa: Deuterostomia: Phylum Chordata: Subphylum Vertebrata: Superclass Cyclostomi
The living jawless vertebrate lineages (Superclass Cyclostom) include hagfish and lampreys. These animals are important because they resemble early branches of vertebrate evolution in some key ways, especially in their lack of jaws. However, they should not be described as unchanged “primitive” animals. Hagfish and lampreys are modern organisms with specialized adaptations of their own. Hagfish are marine animals known for producing large amounts of slime when threatened. They have a skull but lack true vertebrae, which is why they are sometimes discussed as craniates rather than typical vertebrates. Hagfish feed mostly as scavengers and predators of small or weakened animals. They use tooth-like keratin structures to rasp tissue from food. Lampreys have a more vertebrate-like body plan than hagfish, including rudimentary vertebral elements made of cartilage. Many lampreys are parasitic as adults, attaching to fishes with a round sucker-like mouth and feeding on blood or body fluids. Other lamprey species are nonparasitic as adults and do not feed after metamorphosis. Lampreys also have a larval stage that lives in sediment and suspension feeds, which gives scientists insight into early chordate and vertebrate feeding strategies. Hagfish and lampreys show that jaws are not required for a vertebrate-like body plan. However, the later evolution of jaws dramatically changed vertebrate feeding and helped produce many new ecological roles.
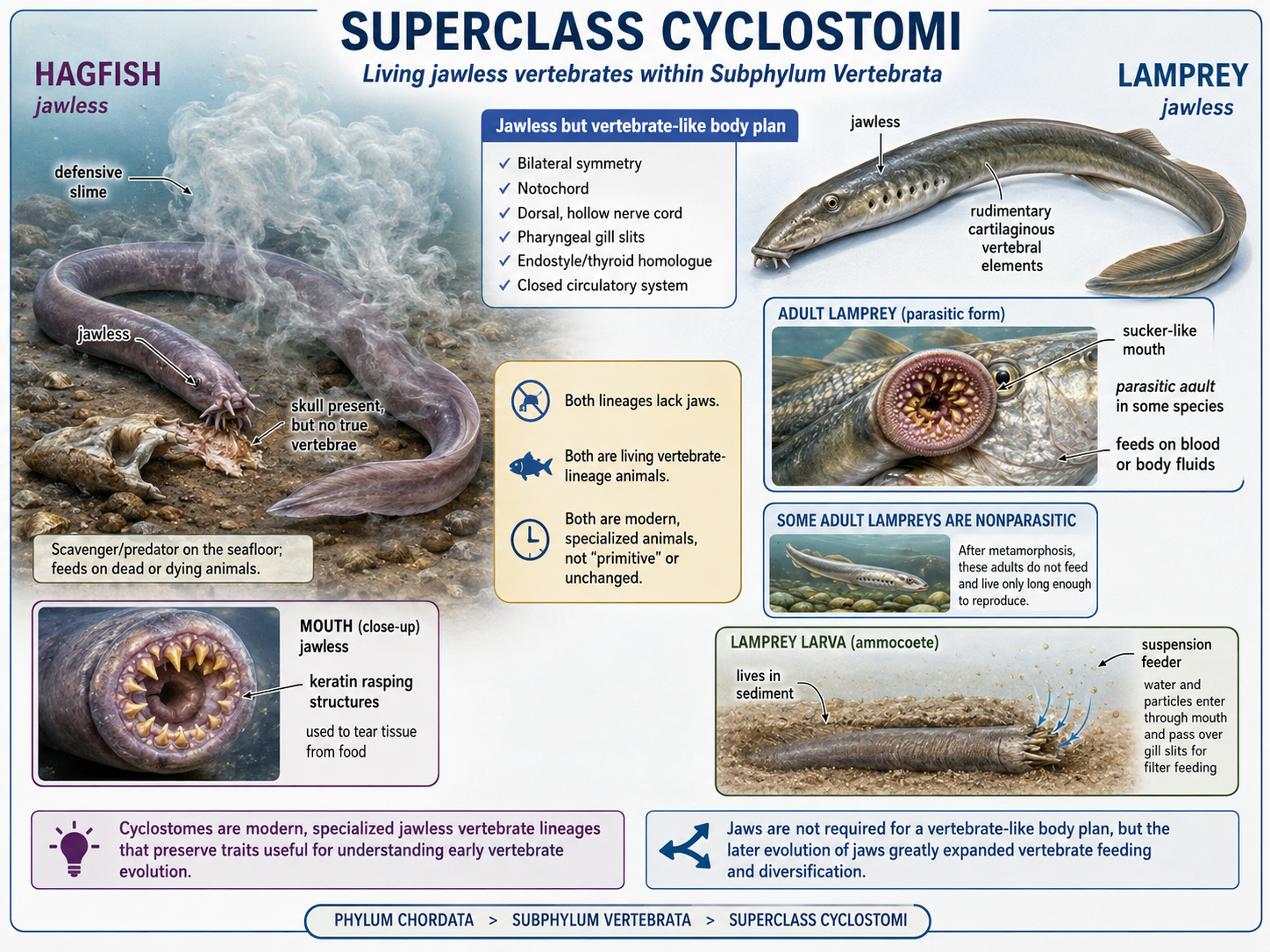
Figure 43. Superclass Cyclostomi. Cyclostomes include hagfish and lampreys, modern jawless vertebrate-lineage animals that help illuminate early vertebrate evolution. Hagfish produce defensive slime, have a skull but lack true vertebrae, and use keratin rasping structures to tear food. Lampreys have rudimentary cartilaginous vertebral elements and may be parasitic or nonparasitic as adults. Their sediment-dwelling larvae suspension feed. Together, cyclostomes show that vertebrate-like body plans evolved before jaws.
Superclass Gnathostomata: Jawed Vertebrates
Kingdom Animalia: Eumetazoa: Bilateria: Nephrozoa: Deuterostomia: Phylum Chordata: Subphylum Vertebrata: Superclass Gnathostomata
Most living vertebrates are jawed vertebrates (Superclass Gnathostomata). Jaws evolved from skeletal supports associated with the pharyngeal arches near the mouth and gill region. This innovation transformed vertebrate evolution. With jaws, vertebrates could bite, grasp, crush, scrape, and process food much more effectively than jawless forms. Jaws were accompanied by other important changes, including paired fins and more complex sensory and nervous systems. Paired fins improved stability and steering during movement, allowing more precise swimming. Later, in one lineage of lobe-finned fishes, paired fins gave rise to limbs capable of supporting movement in shallow water and eventually on land.

Figure 44. Superclass Gnathostomata. Most living vertebrates are jawed vertebrates, or gnathostomes. Jaws evolved from skeletal supports near the mouth and gill region, allowing vertebrates to bite, grasp, crush, scrape, and process food more effectively. Jawed vertebrates also evolved paired fins and more complex sensory and nervous systems, improving movement and control. In one lineage of lobe-finned fishes, paired fins later gave rise to limbs used for movement in shallow water and eventually on land.
Class Chondrichthyes: Sharks, Rays, Skates, and Chimaeras
Kingdom Animalia: Eumetazoa: Bilateria: Nephrozoa: Deuterostomia: Phylum Chordata: Subphylum Vertebrata: Superclass Gnathostomata: Class Chondrichthyes
Class Chondrichthyes includes sharks, rays, skates, and chimaeras. These animals are jawed vertebrates with cartilaginous skeletons. Cartilage is lighter and more flexible than bone, but it can still provide strong support. Many cartilaginous fishes also have mineralized tissues that strengthen parts of the skeleton. Sharks are among the most familiar cartilaginous fishes and are often active predators. Many have streamlined bodies, powerful muscles, large tail fins, and sharp teeth that are continually replaced throughout life. These traits make many sharks effective hunters. However, not all sharks are predators of large animals. Some, such as whale sharks and basking sharks, are enormous suspension feeders that strain plankton from the water. Rays and skates are cartilaginous fishes with flattened bodies. Many live near the seafloor, where they feed on mollusks, crustaceans, worms, and small fishes. Their enlarged pectoral fins form broad, winglike surfaces used in swimming. Some rays, such as manta rays, are active open-water swimmers that filter small organisms from the water. Chimaeras, also called ghost sharks, are less familiar cartilaginous fishes that mostly live in deep marine waters. They have distinctive skull and jaw structures, with the upper jaw fused to the skull. Many have large eyes, tapering tails, and tooth plates used to crush hard-shelled prey and other small bottom-dwelling animals. Cartilaginous fishes also have highly developed sensory systems. Many can detect vibrations, chemicals, and weak electrical fields produced by other animals. These senses make them effective predators and scavengers in marine environments. Although sharks are sometimes portrayed only as dangerous predators, they play important ecological roles by helping regulate marine food webs.

Figure 45. Class Chondrichthyes. Chondrichthyans are jawed vertebrates with cartilaginous skeletons and include sharks, rays and skates, and chimaeras. Sharks are often active predators, though some, such as whale sharks and basking sharks, are suspension feeders. Rays and skates have flattened bodies and enlarged pectoral fins and often feed on bottom-dwelling animals, while manta rays filter small organisms in open water. Chimaeras are deep-marine fishes with large eyes, tooth plates, and an upper jaw fused to the skull.
Clade Osteichthyes: The Evolution of Bone
Kingdom Animalia: Eumetazoa: Bilateria: Nephrozoa: Deuterostomia: Phylum Chordata: Subphylum Vertebrata: Superclass Gnathostomata: Clade Osteichthyes
The other major living lineage of jawed vertebrates includes the bony vertebrates (Clade Osteichthyes). Unlike cartilaginous fishes, which have skeletons made mostly of cartilage, bony vertebrates have skeletons made partly or mostly of bone. Bone is living tissue. It can grow, repair itself, store minerals, and provide strong attachment points for muscles. This stronger internal skeleton became an important foundation for the later evolution of larger and more powerful vertebrate bodies. Bony vertebrates of Clade Osteichthyes include ray-finned fishes, lobe-finned fishes, and tetrapods. Tetrapods are the vertebrates with limbs, including amphibians, reptiles, birds, and mammals. This means that land vertebrates are part of the larger bony vertebrate lineage. From an evolutionary perspective, amphibians, reptiles, birds, and mammals are descended from ancient lobe-finned bony fishes.
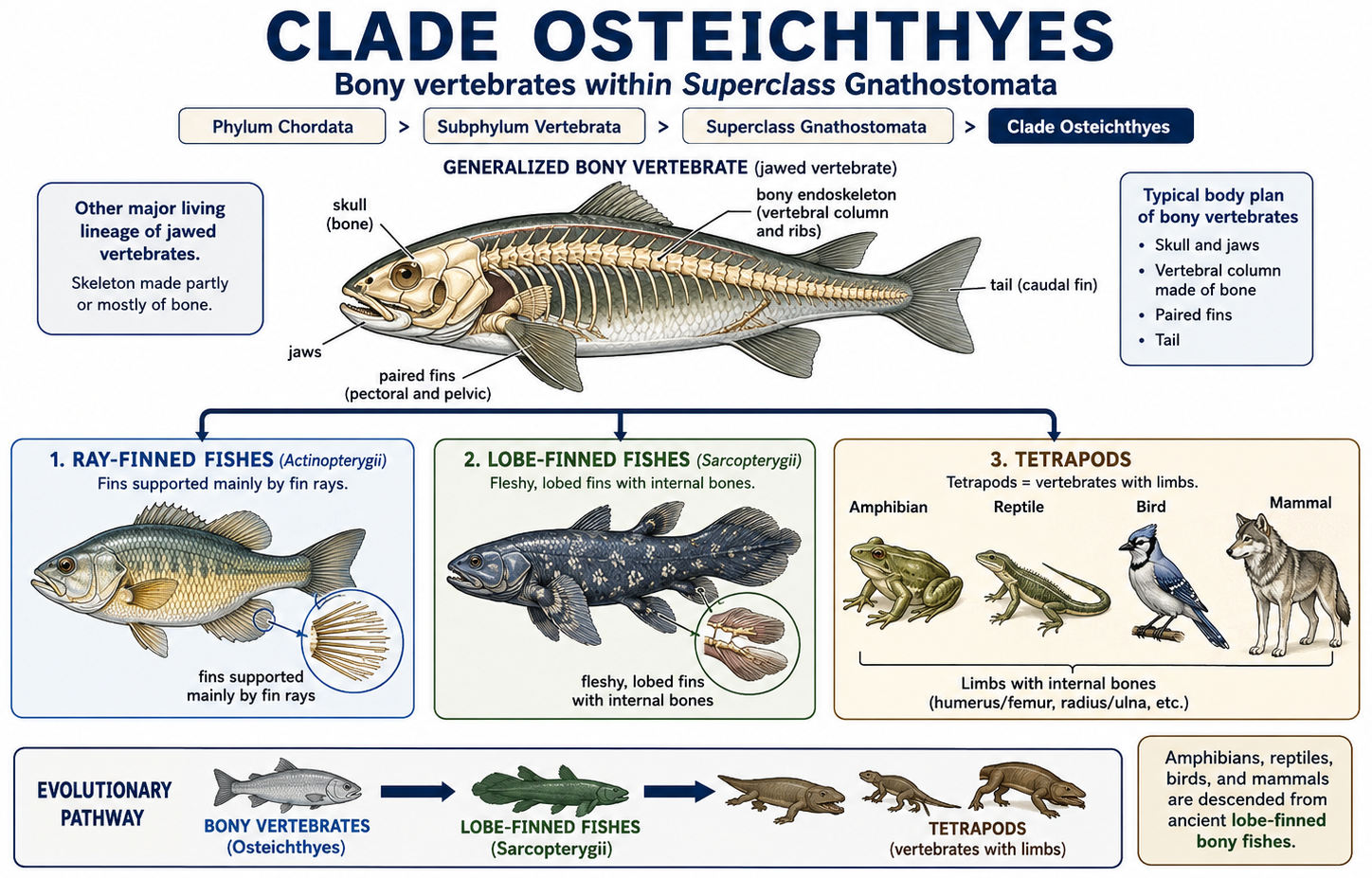
Figure 45. Clade Osteichthyes. Osteichthyans are jawed vertebrates with skeletons made partly or mostly of bone. This clade includes ray-finned fishes, lobe-finned fishes, and tetrapods. Ray-finned fishes have fins supported by fin rays, while lobe-finned fishes have fleshy fins with internal bones. Tetrapods—including amphibians, reptiles, birds, and mammals—evolved from ancient lobe-finned bony fishes and are nested within the larger bony vertebrate lineage.
Class Actinopterygii: Ray-Finned Fishes
Kingdom Animalia: Eumetazoa: Bilateria: Nephrozoa: Deuterostomia: Phylum Chordata: Subphylum Vertebrata: Superclass Gnathostomata: Clade Osteichthyes: Class Actinopterygii
Class Actinopterygii includes the ray-finned fishes, the most diverse group of living vertebrates. Most of the fishes people encounter, such as salmon, trout, tuna, bass, goldfish, seahorses, eels, and clownfish, are ray-finned fishes. Ray-finned fishes have fins supported by thin bony rods called rays. These fins can be moved with great precision, allowing ray-finned fishes to maneuver through coral reefs, rivers, lakes, kelp forests, deep oceans, and many other aquatic habitats. Their body shapes are extremely diverse. Some are fast open-water predators, some are bottom-dwellers, some live in caves, and some have evolved unusual forms such as the upright body of seahorses or the flattened body of flounders. In ray-finned fishes, the swim bladder emerged as an important evolutionary adaptation for buoyancy control. This gas-filled organ allowed fishes to maintain position in the water column with less energy, supporting more efficient swimming and greater ecological diversification. Evolutionarily, the swim bladder is related to air-filled organs such as lungs, reflecting an ancient connection between buoyancy and breathing structures in early bony fishes. The success of ray-finned fishes is tied to their flexible body plan, efficient gills, buoyancy control, jaws, and diverse feeding adaptations. They may feed on plankton, algae, insects, crustaceans, mollusks, other fishes, or detritus. Because they occupy so many ecological roles, ray-finned fishes are central to freshwater and marine ecosystems.

Figure 46. Class Actinopterygii. Ray-finned fishes are the most diverse living vertebrates and include most familiar fishes. Their fins are supported by fin rays, which allow precise movement, and many use a swim bladder for buoyancy control. Their wide variety of forms—including open-water swimmers, bottom-dwellers, eels, seahorses, and reef fishes—reflects their major ecological success in freshwater and marine ecosystems.
Class Sarcopterygii (Lobe-Finned Fishes)
Kingdom Animalia: Eumetazoa: Bilateria: Nephrozoa: Deuterostomia: Phylum Chordata: Subphylum Vertebrata: Superclass Gnathostomata: Clade Osteichthyes: Class Sarcopterygii
Class Sarcopterygii includes the lobe-finned fishes and the tetrapod lineage. Living non-tetrapod sarcopterygians include lungfishes and coelacanths. Lobe-finned fishes differ from ray-finned fishes because their paired fins are supported by fleshy, muscular lobes containing internal bones. These bones are evolutionarily significant because they resemble the pattern that later gave rise to tetrapod limbs. Lungfishes are especially important for understanding the transition from water to land. They have lungs or lung-like organs and can breathe air. Some lungfishes live in habitats that may become low in oxygen or dry seasonally. During droughts, certain lungfish can survive by burrowing into mud and entering a dormant state. Their ability to breathe air does not mean they are “half fish, half amphibian,” but it does show how air breathing and strong paired fins could have been useful in shallow-water environments. Coelacanths are another living lineage of lobe-finned fishes. They were known from fossils before living specimens were discovered in the twentieth century. Because of this, coelacanths are often called “living fossils,” although that phrase can be misleading. Modern coelacanths are not identical to their ancient relatives. They are living species that have continued evolving, even if their overall body form resembles some fossil forms. The tetrapods evolved from within the lobe-finned fish lineage. This means that amphibians, reptiles, birds, and mammals are deeply connected to sarcopterygian ancestry. The bones of our arms, wrists, hands, legs, ankles, and feet have evolutionary roots in the paired fins of ancient lobe-finned fishes.

Figure 47. Class Sarcopterygii. Sarcopterygians include lungfishes, coelacanths, and the lineage that gave rise to tetrapods. Their paired fins are fleshy, muscular lobes supported by internal bones, linking lobe-finned fish anatomy to the later evolution of limbs. Lungfishes can breathe air and survive drought in mud burrows, while coelacanths are distinctive deep-water forms first known from fossils. This group shows that amphibians, reptiles, birds, and mammals evolved from within the lobe-finned fish lineage.
Clade Tetrapoda: Move onto Land
Kingdom Animalia: Eumetazoa: Bilateria: Nephrozoa: Deuterostomia: Phylum Chordata: Subphylum Vertebrata: Superclass Gnathostomata: Clade Osteichthyes: Class Sarcopterygii: Clade Tetrapoda
Clade Tetrapoda includes the vertebrate lineage that gave rise to amphibians, reptiles, birds, and mammals. The name Tetrapoda means “four feet,” although not all living tetrapods have four limbs. Some groups have reduced, lost, or modified limbs, but they are still tetrapods because they descend from four-limbed ancestors. Tetrapods evolved from within the lobe-finned fish lineage. The earliest tetrapods lived in shallow aquatic environments and were not fully land-dwelling animals. Their evolution involved major changes to the skeleton, muscles, lungs, skull, neck, ribs, vertebral column, and sensory systems. The vertebral column and ribs became more important for resisting gravity, while lungs became increasingly important for breathing air. Changes in the skull and neck allowed the head to move more independently, and sensory systems had to work in air instead of water. Early tetrapods still depended on aquatic habitats for reproduction. Like many modern amphibians, they likely produced eggs that were vulnerable to drying out. Full independence from aquatic reproduction evolved later with the amniotic egg, which protected embryos from drying. This innovation gave rise to the amniotes, the lineage that includes reptiles, birds, and mammals. Tetrapod evolution was not a sudden leap from fish to land animals. It was a gradual accumulation of traits that helped vertebrates with lobe-finned ancestry move, breathe, sense, and reproduce in increasingly terrestrial environments.
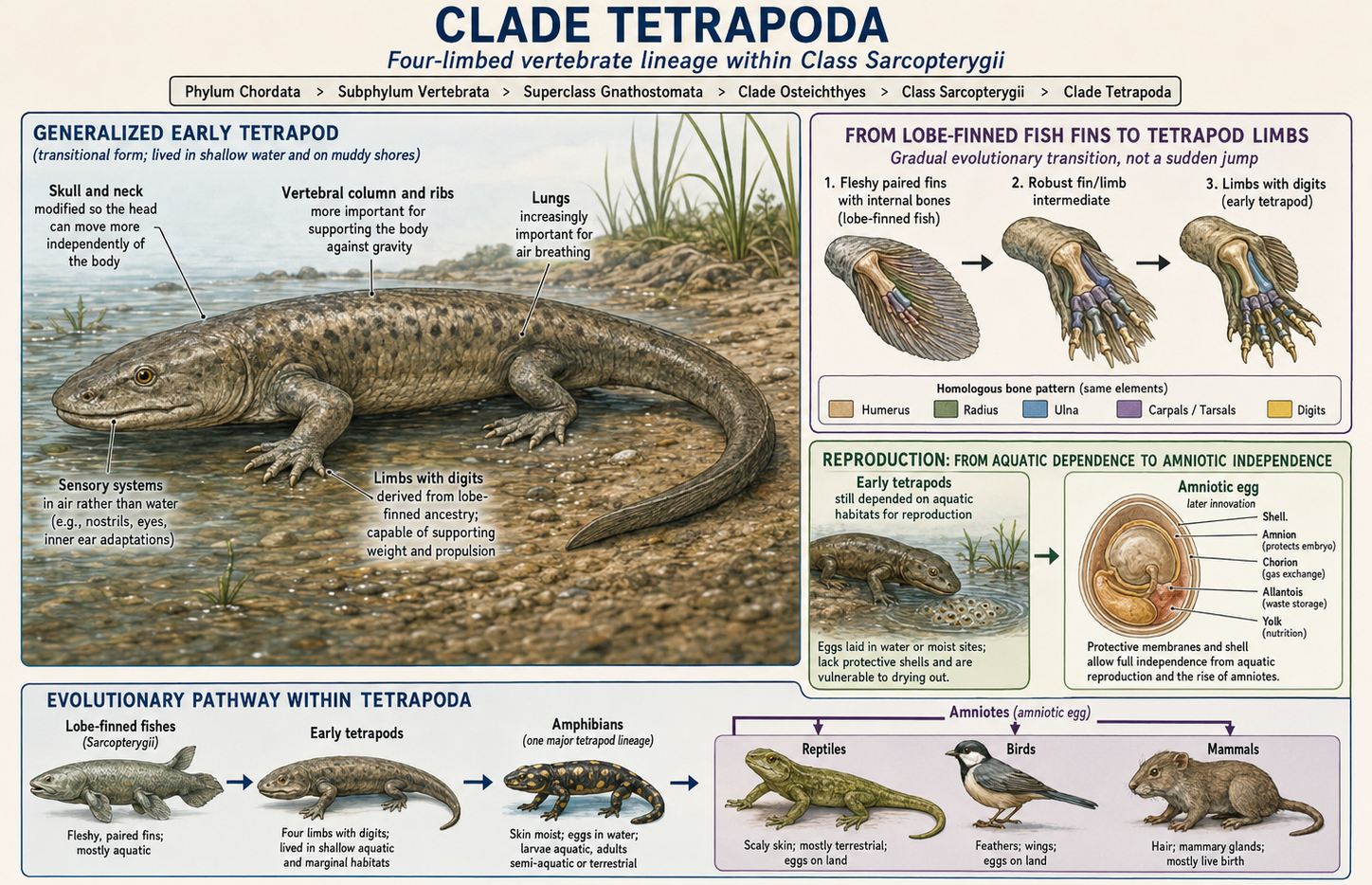
Figure 48. Clade Tetrapod: Early tetrapods and the move onto land. Tetrapods evolved from within the lobe-finned fish lineage and gave rise to amphibians, reptiles, birds, and mammals. Early tetrapods lived in shallow-water habitats and show the transition from fleshy fins to limbs with digits, along with stronger vertebral columns and ribs, increased use of lungs, and changes in the skull and neck that allowed the head to move more independently. Early tetrapods still depended on water for reproduction, but the later evolution of the amniotic egg allowed full independence from aquatic breeding in the amniote lineage.
Class Amphibia: Amphibians (Frogs, Toads, Salamanders, Newts, and Caecilians)
Kingdom Animalia: Eumetazoa: Bilateria: Nephrozoa: Deuterostomia: Phylum Chordata: Subphylum Vertebrata: Superclass Gnathostomata: Clade Osteichthyes: Class Sarcopterygii: Clade Tetrapoda: Class Amphibia
Class Amphibia includes frogs, toads, salamanders, newts, and caecilians. Amphibians are tetrapods, but most remain closely tied to moist environments. Many amphibians have thin, permeable skin that can exchange gases and water with the environment. This skin makes them vulnerable to drying out, pollution, and disease, but it also allows many species to supplement lung breathing with cutaneous respiration, or breathing through the skin. Amphibian life cycles are diverse. Many frogs and salamanders lay eggs in water, and their larvae hatch as aquatic forms that later metamorphose into adults. Tadpoles, for example, are the aquatic larvae of frogs and toads. However, not all amphibians follow this pattern. Some lay eggs on land in moist places, some show direct development without a free-swimming larval stage, and some give birth to live young. Amphibians are usually carnivorous as adults, feeding on insects, worms, small crustaceans, and other animals. Their larvae may be herbivorous, omnivorous, or carnivorous depending on the species. Frogs and toads are often specialized for jumping, salamanders usually have a more elongated body and tail, and caecilians are limbless burrowing amphibians that superficially resemble worms or snakes. Caecilians show why it is important not to define amphibians simply as “four-legged animals.” They belong to Tetrapoda by ancestry, even though their limbs have been lost.
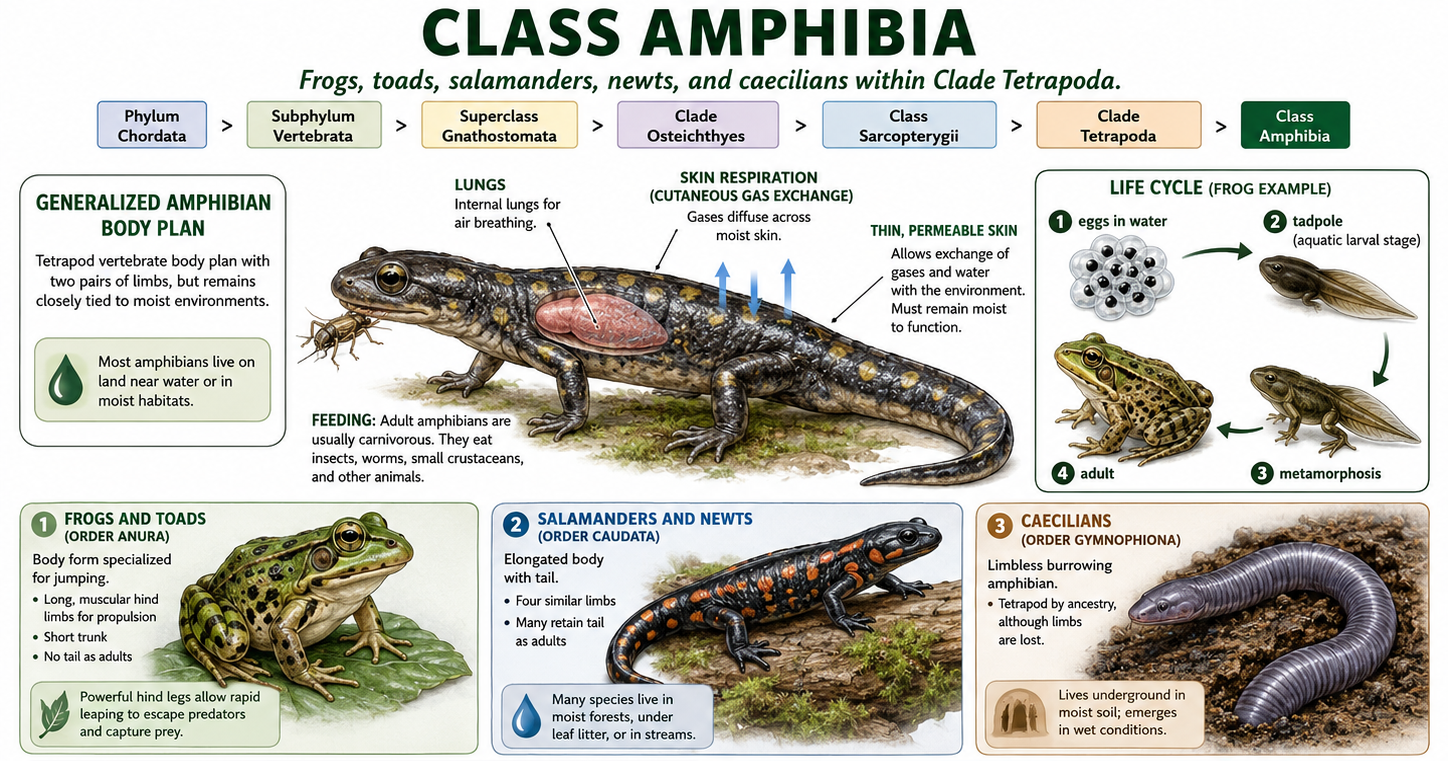
Figure 49. Class Amphibia. Moist-skinned tetrapods with diverse life cycles. Amphibians include frogs, toads, salamanders, newts, and caecilians. Although they are tetrapods, most remain tied to moist habitats because their thin, permeable skin can exchange water and gases, allowing cutaneous respiration but also increasing vulnerability to drying, pollution, and disease. Many species lay eggs in water and pass through aquatic larvae, such as tadpoles, before metamorphosis, while others develop directly or give birth to live young. Adult amphibians are usually carnivorous. Caecilians show that tetrapods are defined by ancestry, not simply by having four limbs.
Clade Amniota: Amniotes (Reptiles, Birds and Mammals)
Kingdom Animalia: Eumetazoa: Bilateria: Nephrozoa: Deuterostomia: Phylum Chordata: Subphylum Vertebrata: Superclass Gnathostomata: Clade Osteichthyes: Class Sarcopterygii: Clade Tetrapoda: Clade Amniota
Clade Amniota includes reptiles, birds, mammals, and their extinct relatives. Amniotes are tetrapods, but unlike most amphibians, they are not tied to open water for reproduction. The key innovation of this group is the amniotic egg, which protects the developing embryo with specialized membranes. These membranes create a controlled fluid environment around the embryo, allowing development to occur on land. The amniotic egg contains several important extra-embryonic membranes. The amnion surrounds the embryo with fluid and helps prevent drying out. The yolk sac provides nutrients. The chorion helps with gas exchange between the embryo and the outside environment. The allantois stores nitrogenous wastes and also contributes to gas exchange in many species. In reptiles and birds, the amniotic egg is usually enclosed by a leathery or hard shell. In most mammals, the embryo develops inside the mother, and some of these same membranes help form the placenta. The evolution of the amniotic egg was one of the most important transitions in vertebrate history. It allowed vertebrates to reproduce away from open water and helped amniotes diversify into drier terrestrial habitats. Amniotes later split into two major evolutionary lineages. Clade Sauropsida includes reptiles, birds, and their extinct relatives. Clade Synapsida includes mammals and their extinct relatives. This split produced the two main branches of land vertebrates that dominate many terrestrial ecosystems today.
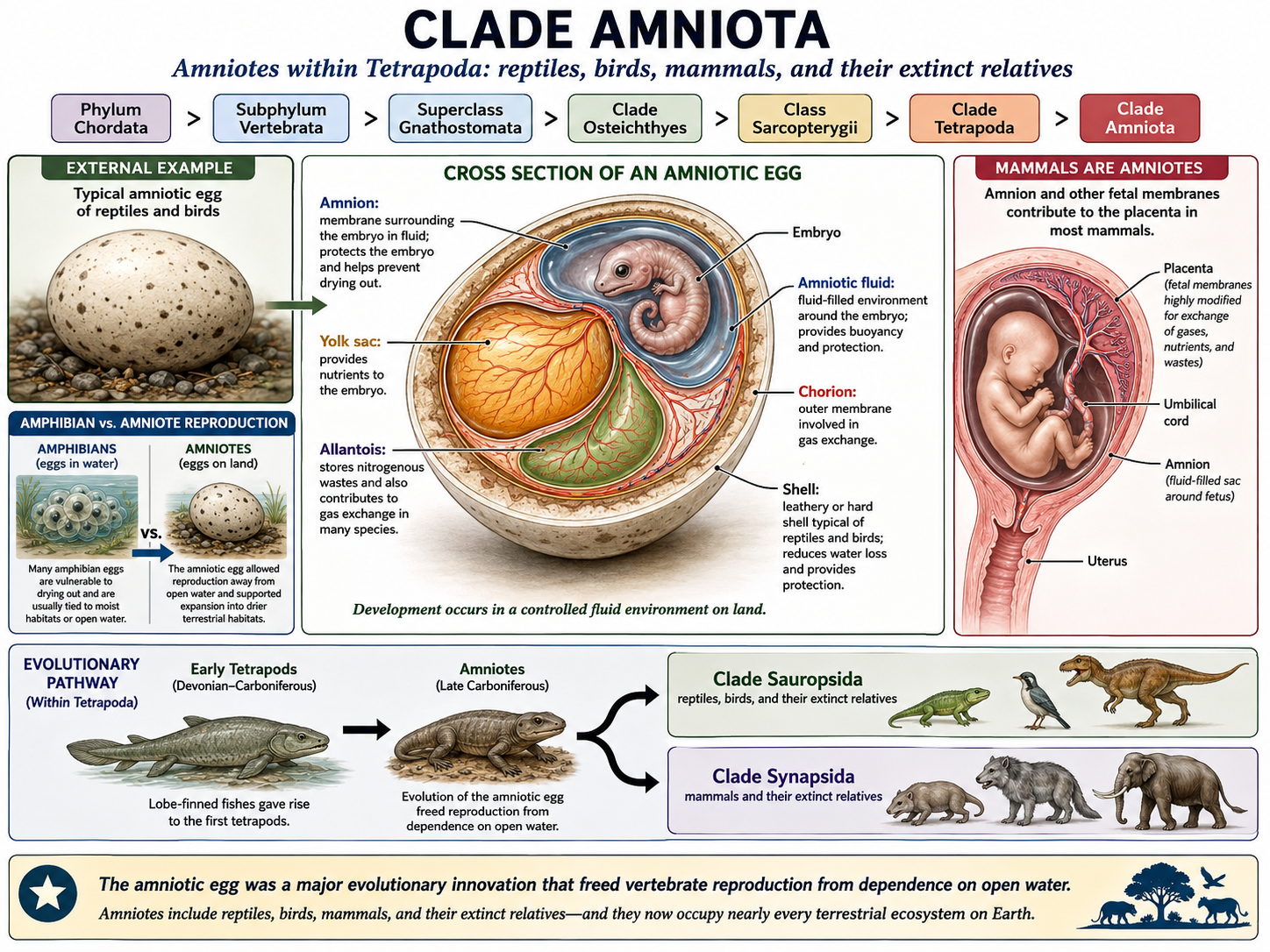
Figure 50. Clade Amniota. The amniotic egg and reproduction on land. Amniotes include reptiles, birds, mammals, and their extinct relatives. Their defining innovation is the amniotic egg, whose extra-embryonic membranes—amnion, yolk sac, chorion, and allantois—protect the embryo, provide nutrients, support gas exchange, and help manage wastes in a controlled fluid environment. This adaptation freed reproduction from open water, unlike the eggs of most amphibians, and helped amniotes expand into drier terrestrial habitats. Amniotes later split into Sauropsida and Synapsida, the lineages leading to reptiles and birds, and to mammals.
Class Reptilia: Turtles, Lizards, Snakes, Tuatara, Crocodilians, and Birds
Kingdom Animalia: Eumetazoa: Bilateria: Nephrozoa: Deuterostomia: Phylum Chordata: Subphylum Vertebrata: Superclass Gnathostomata: Clade Osteichthyes: Sarcopterygii: Clade Tetrapoda: Clade Amniota: Clade Sauropsida: Class Reptilia
Class Reptilia, used here in its modern evolutionary sense, includes turtles, lizards, snakes, tuatara, crocodilians, birds, and their extinct relatives. Birds are included because they evolved from theropod dinosaurs, which were reptiles. If birds are left out, reptiles become an incomplete evolutionary group because the group would exclude some descendants of a reptile ancestor. For this reason, many biologists treat birds as living reptiles. Reptiles are amniotes, and many of their features reflect adaptation to life on land. Most have skin covered with keratinized scales, plates, or feathers that help protect the body and reduce water loss. Reptiles breathe with lungs throughout life, and most reproduce using amniotic eggs. Modern reptiles are highly diverse. Turtles have protective shells formed partly from the skeleton. Lizards show many different feeding and movement strategies. Snakes are limbless reptiles with flexible skulls and specialized feeding adaptations. Tuatara are the only living members of an old reptile lineage. Crocodilians are powerful semi-aquatic predators with strong jaws and parental care. Birds are feathered reptiles that evolved from dinosaurs and became one of the most diverse groups of living land vertebrates. Most nonavian reptiles are ectothermic, meaning they rely heavily on external heat sources to regulate body temperature. Birds are endothermic, meaning they produce most of their body heat through metabolism. This difference is important, but it does not remove birds from Reptilia. Instead, it shows that major evolutionary changes occurred within the reptile lineage. In modern evolutionary classification, Reptilia includes both familiar nonavian reptiles and birds, along with extinct groups such as nonavian dinosaurs and pterosaurs.
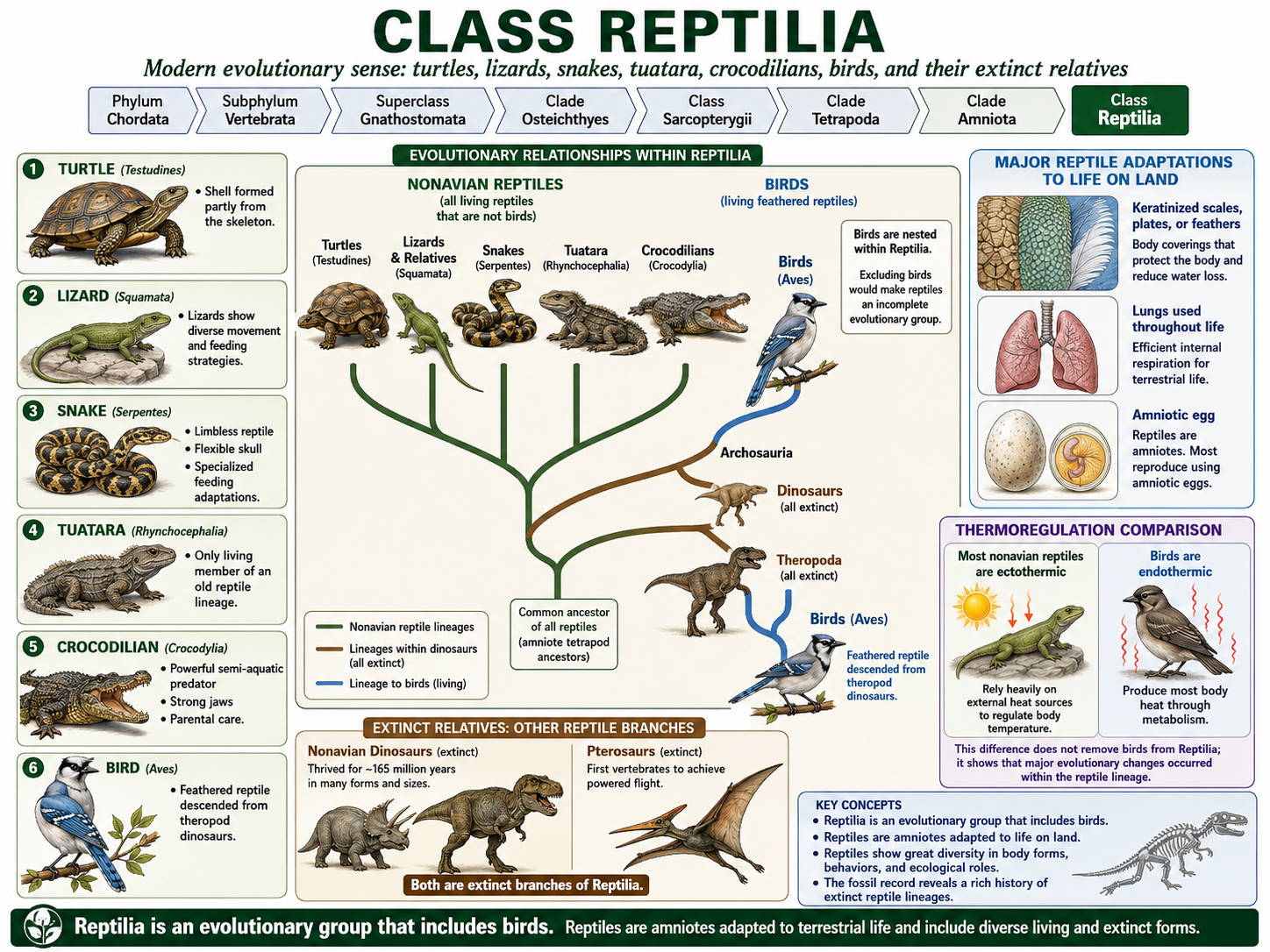
Figure. Class Reptilia. Reptiles include turtles, lizards, snakes, tuatara, crocodilians, birds, and their extinct relatives. In evolutionary terms, birds are reptiles because they descended from theropod dinosaurs, so excluding birds would make Reptilia incomplete. Reptiles are amniotes with adaptations for life on land, including keratinized scales, plates, or feathers, lungs used throughout life, and typically an amniotic egg. Living reptiles show wide diversity in body form and biology, from mostly ectothermic nonavian reptiles to endothermic birds, as well as extinct groups such as nonavian dinosaurs and pterosaurs.
Order Testudines: Turtles and Tortoises
Kingdom Animalia: Eumetazoa: Bilateria: Nephrozoa: Deuterostomia: Phylum Chordata: Subphylum Vertebrata: Superclass Gnathostomata: Clade Osteichthyes: Sarcopterygii: Clade Tetrapoda: Clade Amniota: Clade Sauropsida: Class Reptilia: Order Testudines
Order Testudines includes turtles, tortoises, and terrapins. These reptiles are recognized by their shell, which is made of expanded ribs, vertebrae, and bony plates. The upper part of the shell is the carapace, and the lower part is the plastron. The shell is not an external object that the animal can leave behind; it is part of the skeleton. Turtles lack teeth and instead have a keratinized beak. Their diets vary widely. Some are herbivores, some are carnivores, and some are omnivores. Tortoises are mostly terrestrial, while many turtles are aquatic or semi-aquatic. Sea turtles are highly adapted for marine life, with limbs modified into flippers.

Figure 52. Order Testudines. Turtles, tortoises, and terrapins. Testudines are reptiles recognized by a protective shell made from expanded ribs, vertebrae, and bony plates. The upper shell is the carapace, and the lower shell is the plastron; the shell is part of the skeleton, not something the animal can leave behind. Turtles lack teeth and use a keratinized beak to feed on plants, animals, or both. Tortoises are mostly terrestrial, many turtles are aquatic or semi-aquatic, and sea turtles have flipper-like limbs adapted for marine life.
Order Squamata: Lizards and Snakes
Kingdom Animalia: Eumetazoa: Bilateria: Nephrozoa: Deuterostomia: Phylum Chordata: Subphylum Vertebrata: Superclass Gnathostomata: Clade Osteichthyes: Sarcopterygii: Clade Tetrapoda: Clade Amniota: Clade Sauropsida: Class Reptilia: Order Squamata
Order Squamata includes lizards, snakes, and amphisbaenians. Squamates are the most diverse living reptile order. They have scaly skin and a flexible skull structure that allows many species to capture and swallow large or awkward prey. Lizards usually have legs, external ear openings, and movable eyelids, although there are exceptions. Some lizard lineages have reduced limbs or no limbs at all. Snakes are limbless squamates that evolved from lizard-like ancestors. They have elongated bodies, no external ear openings, and highly kinetic skulls. Many snakes use constriction or venom to subdue prey, while others swallow small prey without these specializations. Some snakes retain tiny vestiges of hind limbs, such as pelvic spurs in boas and pythons. These structures are remnants of their evolutionary ancestry and may be used during mating. They are a good example of how evolution modifies existing structures rather than designing organisms from scratch. Amphisbaenians are a specialized group of mostly limbless, burrowing reptiles. They have wormlike bodies, reinforced skulls for digging, reduced or hidden eyes, and feed mainly on small invertebrates underground.
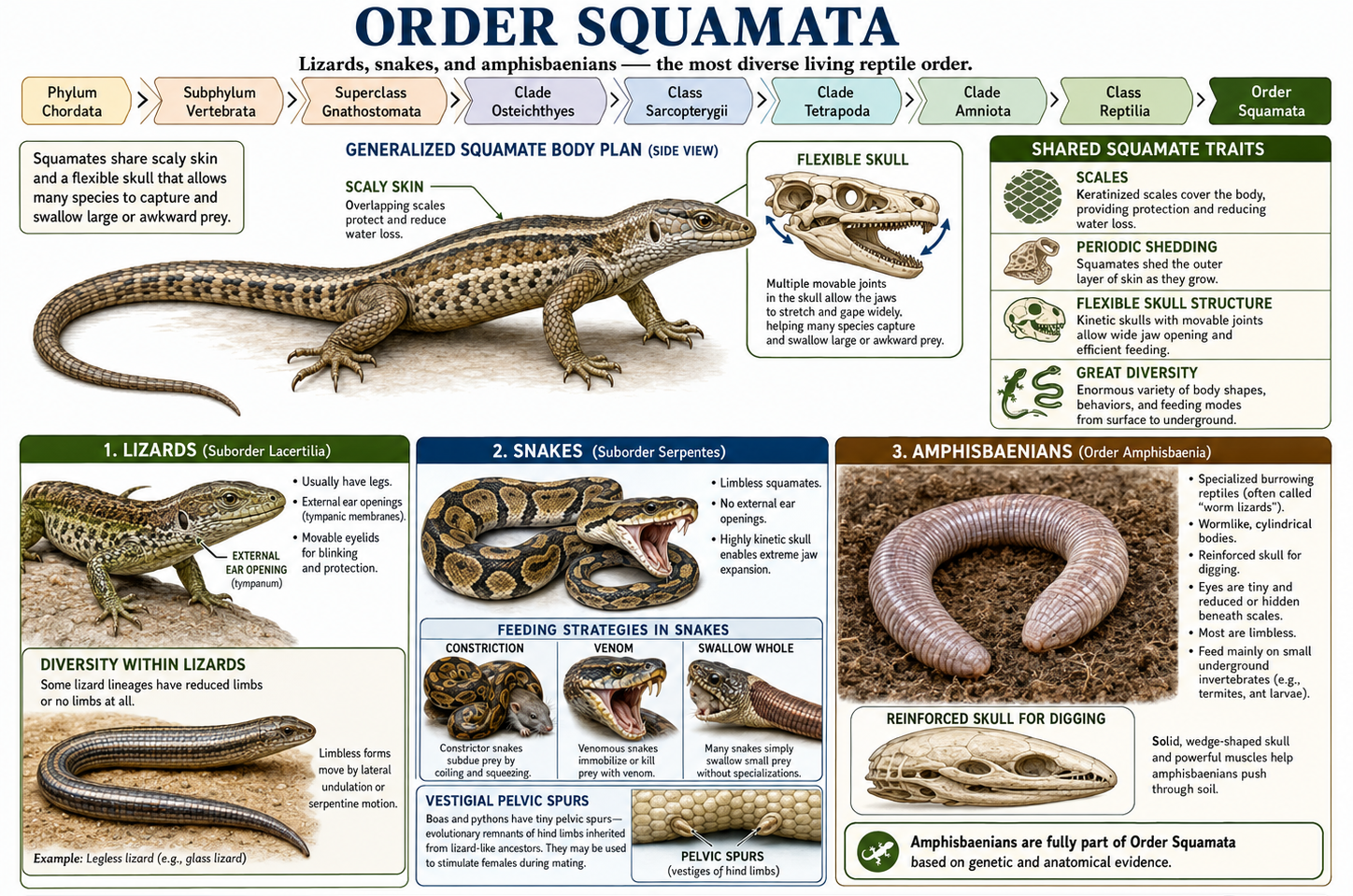
Figure 53. Order Squamata. Lizards, snakes, and amphisbaenians. Squamates are the most diverse living reptile order and include lizards, snakes, and amphisbaenians. They generally have scaly skin and a flexible skull that contributes to feeding versatility. Lizards usually have legs, external ear openings, and movable eyelids, although some lineages have reduced or lost limbs; snakes are limbless squamates with elongated bodies and highly kinetic skulls, and some retain tiny hind limb remnants such as pelvic spurs. Amphisbaenians are specialized burrowing squamates with wormlike bodies, reinforced skulls, and reduced eyes, showing how extensively body form has been modified within the group.
Order Crocodylia: Crocodilians
Kingdom Animalia: Eumetazoa: Bilateria: Nephrozoa: Deuterostomia: Phylum Chordata: Subphylum Vertebrata: Superclass Gnathostomata: Clade Osteichthyes: Sarcopterygii: Clade Tetrapoda: Clade Amniota: Clade Sauropsida: Class Reptilia: Clade Archosauria: Order Crocodylia
Order Crocodylia includes crocodiles, alligators, caimans, and gharials. Crocodilians are in same larger lineage that includes birds and their dinosaur relatives. Because of this shared ancestry, crocodilians are the closest living relatives of birds. Crocodilians are highly adapted for life at the boundary between land and water. Their eyes and nostrils are positioned high on the skull, allowing them to remain mostly submerged while still watching their surroundings and breathing at the water’s surface. Their long, powerful jaws are lined with conical teeth suited for gripping prey rather than chewing it. Strong neck muscles, armored skin, and a muscular tail help them capture prey, defend themselves, and move efficiently through water. Most crocodilians are ambush predators in rivers, lakes, wetlands, estuaries, and coastal habitats. They often wait quietly near the water’s edge, then strike suddenly when fish, birds, mammals, or other animals come within reach. Crocodilians also show complex behaviors. Many build or guard nests, protect eggs, respond to calls from hatchlings, and provide parental care after the young emerge. This level of care shows that reptiles can have more advanced social and reproductive behaviors than many people assume.
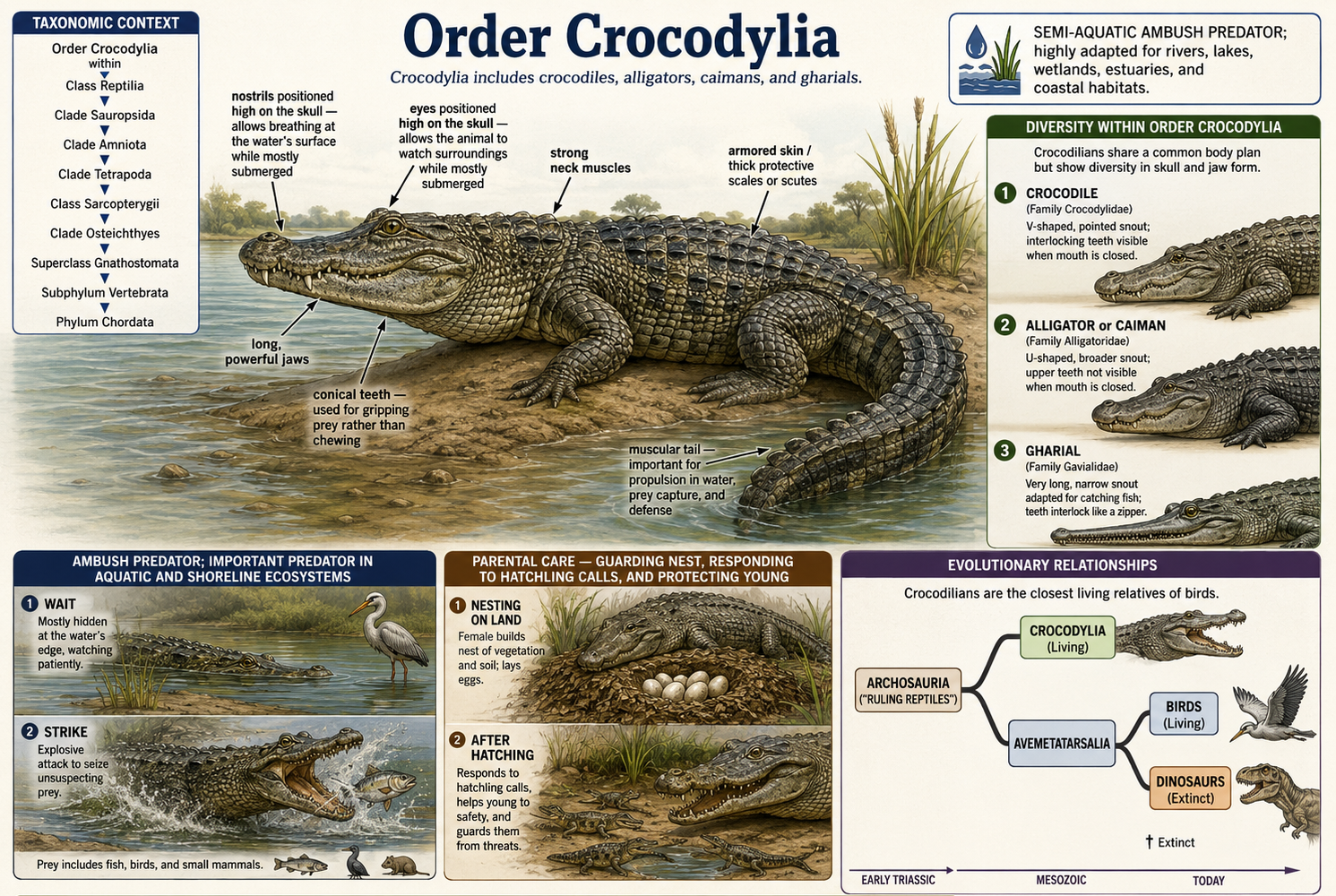
Figure 54. Order Crocodylia. Crocodiles, alligators, caimans, and gharials. Crocodilians are semi-aquatic reptiles and the closest living relatives of birds, reflecting their shared ancestry within the archosaur lineage. Their eyes and nostrils sit high on the skull, allowing them to remain mostly submerged while watching and breathing at the surface. Powerful jaws with conical teeth, armored skin, strong neck muscles, and a muscular tail make them effective ambush predators in rivers, wetlands, estuaries, and coastal habitats. Crocodilians also show complex reproductive behavior, including nest guarding, responses to hatchling calls, and parental care.
Class Aves: Birds
Kingdom Animalia: Eumetazoa: Bilateria: Nephrozoa: Deuterostomia: Phylum Chordata: Subphylum Vertebrata: Superclass Gnathostomata: Clade Osteichthyes: Sarcopterygii: Clade Tetrapoda: Clade Amniota: Clade Sauropsida: Reptilia: Clade Archosauria: Clade Dinosauria: Clade Theropoda: Class Aves
Class Aves includes birds, which are feathered theropod dinosaurs and therefore part of the reptile lineage. Birds are endothermic vertebrates with feathers, beaks, lightweight skeletons, and highly efficient respiratory systems. Most birds are capable of flight, although some, such as ostriches, emus, kiwis, and penguins, are flightless. Feathers are one of the defining traits of birds. They provide insulation, aid in flight, and often play roles in display, camouflage, and communication. Bird bones are often lightweight, and many contain air spaces connected to the respiratory system. Birds also have a unique flow-through lung system with air sacs, allowing highly efficient gas exchange. This respiratory system supports the high metabolic demands of flight. Birds occupy an enormous range of ecological roles. Some are predators, some are seed eaters, some feed on nectar, some filter food from water, and some scavenge. Their beaks, feet, wings, and behaviors are shaped by these ecological roles. Because birds are living dinosaurs, they provide one of the clearest examples of how evolutionary classification can change the way we understand familiar animals.
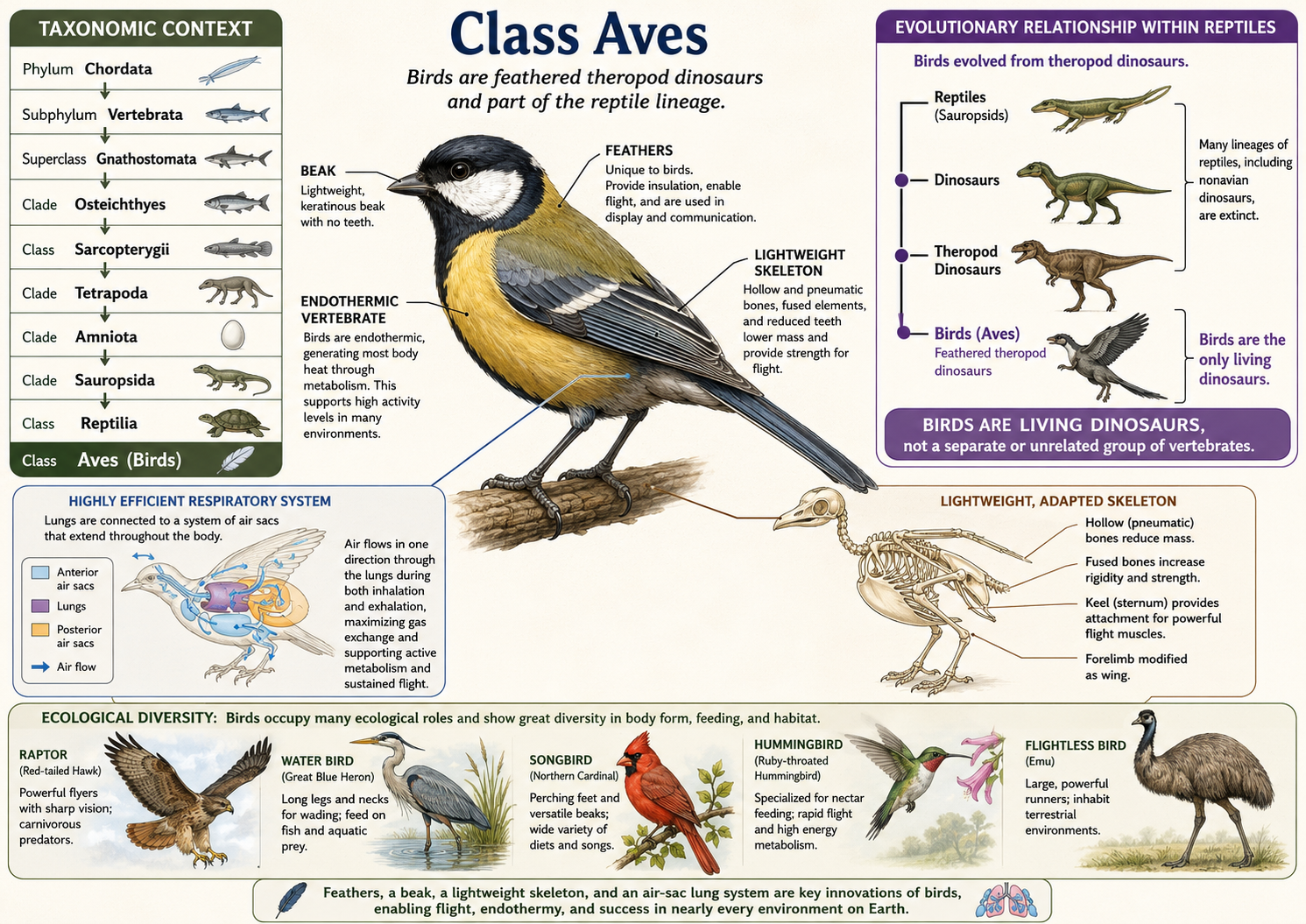
Figure 55. Class Aves. Birds as living feathered reptiles. Birds are feathered theropod dinosaurs and therefore part of Class Reptilia, showing that modern reptiles include more than the traditional nonbird groups. They are distinguished by feathers, a beak, a lightweight skeleton, and a highly efficient air-sac lung system that supports high metabolic activity and, in many species, flight. Birds are also endothermic, generating most of their body heat through metabolism. Their ancestry within the theropod dinosaur lineage makes birds one of the clearest examples of how evolutionary classification changes our understanding of familiar animals.
Class Mammalia: Mammals
Kingdom Animalia: Eumetazoa: Bilateria: Nephrozoa: Deuterostomia: Phylum Chordata: Subphylum Vertebrata: Superclass Gnathostomata: Clade Osteichthyes: Class Sarcopterygii: Clade Tetrapoda: Clade Amniota: Clade Synapsida: Class Mammalia
Class Mammalia includes monotremes, marsupials, and placental mammals. Mammals belong to the amniote lineage separate from reptiles and birds. Mammals are defined by several key traits, including fur, mammary glands, and three middle ear bones. Mammals are also endothermic, meaning they generate body heat through metabolism. Hair helps with insulation, sensory functions, camouflage, and display. Mammary glands allow mothers to nourish young after birth or hatching. The three middle ear bones—the malleus, incus, and stapes—evolved from bones that were part of the jaw joint in earlier synapsid ancestors. This is one of the best examples of how evolution can repurpose existing structures for new functions. Mammals also tend to show high levels of parental care, although the details vary widely among species. Some mammals produce large numbers of poorly developed offspring, while others produce fewer young and invest heavily in each one. Mammalian diversity includes burrowing moles, flying bats, aquatic whales, armored armadillos, egg-laying monotremes, pouch-bearing marsupials, and highly social primates.
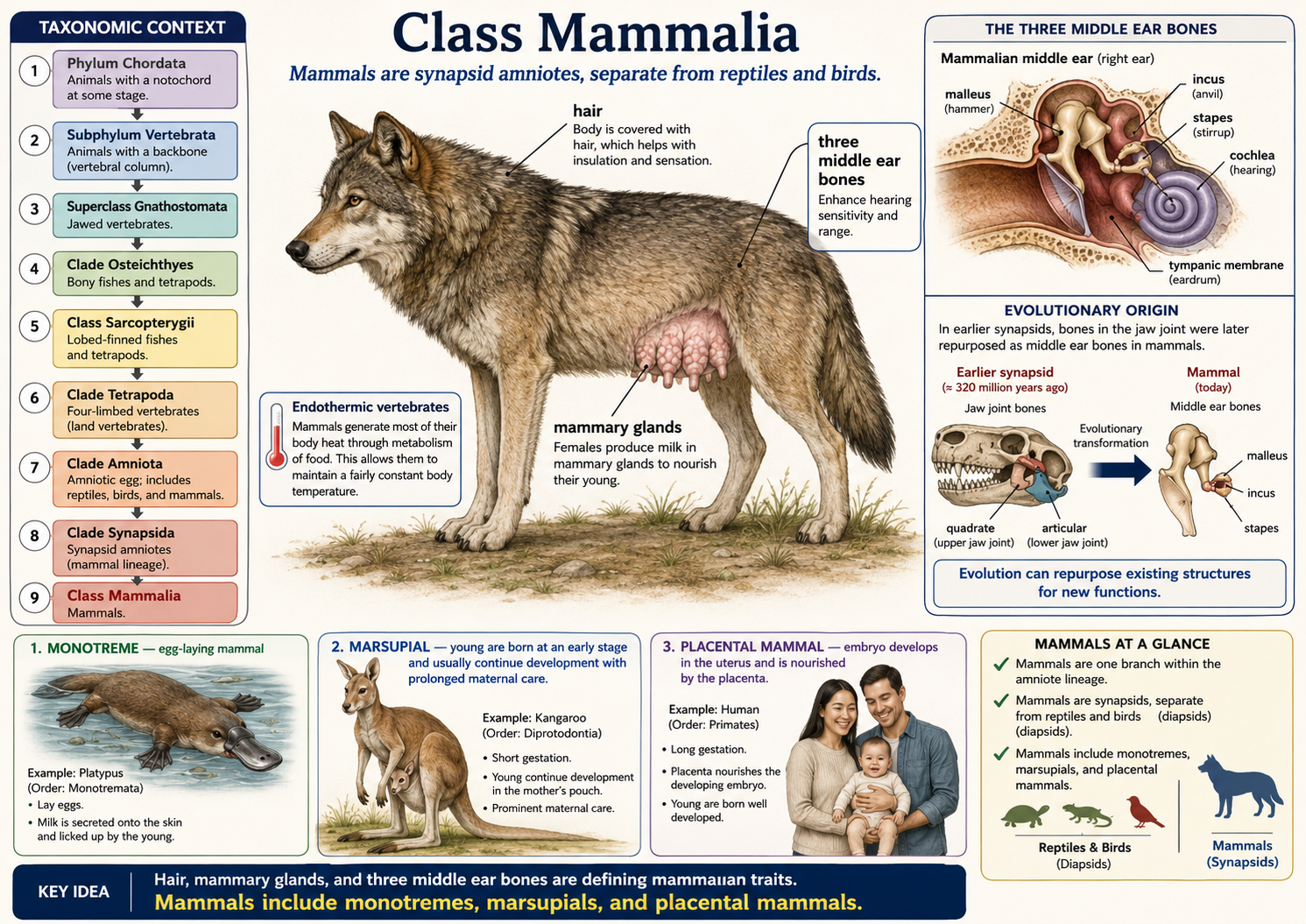
Figure 56. Class Mammalia. Hair, milk production, and specialized ear bones. Mammals are endothermic synapsid amniotes that include monotremes, marsupials, and placental mammals. They are defined by fur or hair, mammary glands, and three middle ear bones—the malleus, incus, and stapes—with hair serving roles in insulation, sensing, camouflage, and display, and mammary glands nourishing young. Monotremes lay eggs, marsupials give birth to highly undeveloped young that continue development with maternal care, and placental mammals nourish embryos through a placenta. Mammals also show great ecological diversity, from burrowers and fliers to swimmers and primates, while retaining the core traits that unite the group.
Infraclass Monotremata: Monotremes
Kingdom Animalia: Eumetazoa: Bilateria: Nephrozoa: Deuterostomia: Phylum Chordata: Subphylum Vertebrata: Superclass Gnathostomata: Clade Osteichthyes: Class Sarcopterygii: Clade Tetrapoda: Clade Amniota: Clade Synapsida: Class Mammalia: Infraclass Monotremata
Infraclass Monotremata includes the egg-laying mammals: the platypus and echidnas. Monotremes are mammals because they have hair and produce milk, but they differ from other living mammals because they lay eggs rather than giving birth to live young. The platypus is a semi-aquatic monotreme with a broad bill, webbed feet, and electroreceptors that help detect prey underwater. Echidnas are terrestrial monotremes with spines, strong claws, and long sticky tongues used to feed on ants and termites. Monotremes do not have nipples; instead, milk is secreted onto specialized areas of skin where the young lap it up. Monotremes are sometimes described as “primitive mammals,” but that wording can be misleading. They retain some ancestral traits, such as egg laying, but they also have specialized adaptations that evolved within their own lineage.
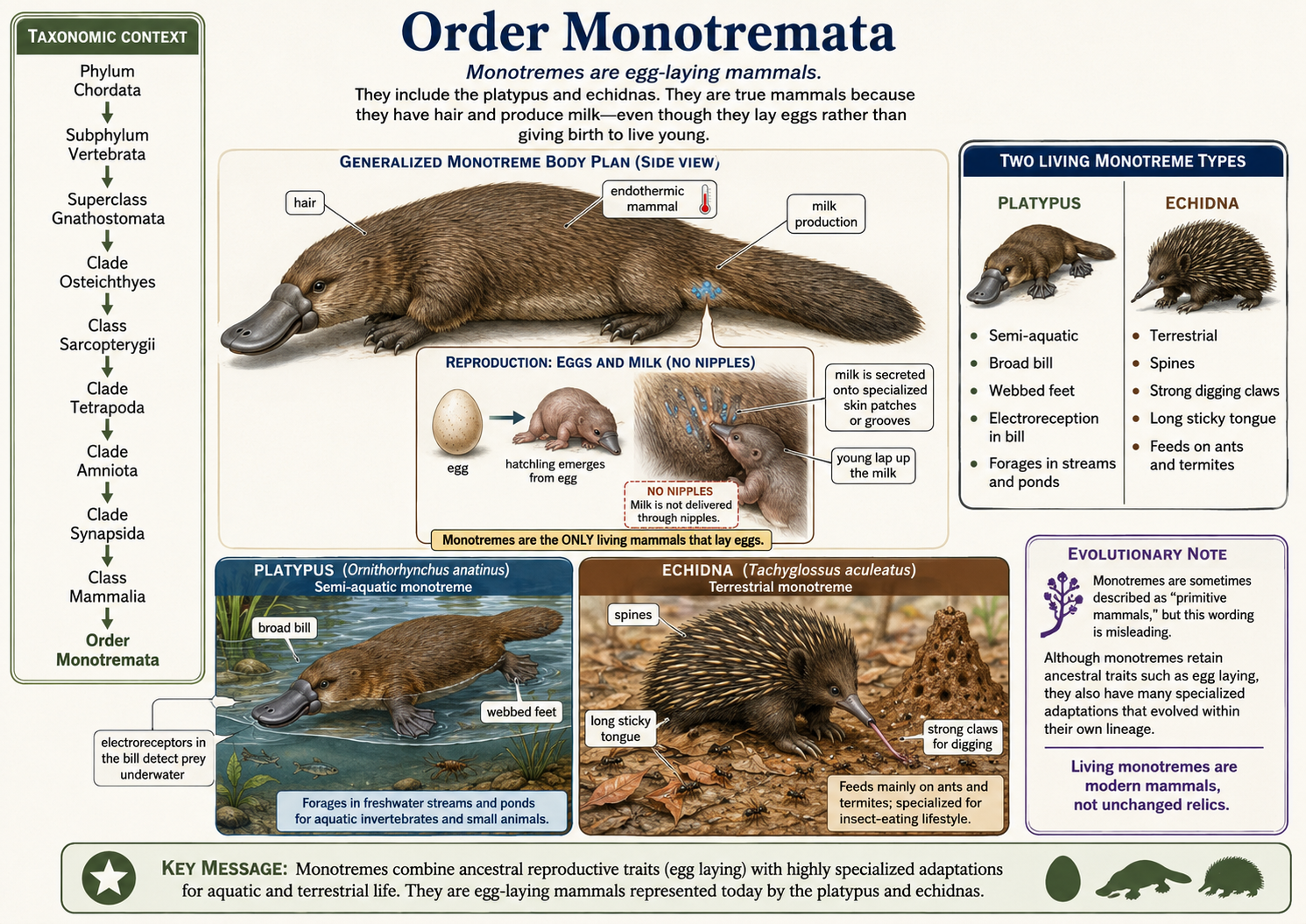
Figure 57. Order Monotremata. Egg-laying mammals. Monotremes are true mammals because they have hair and produce milk, but unlike other living mammals they lay eggs. Living monotremes include the platypus, a semi-aquatic form with a broad bill, webbed feet, and electroreception, and the echidnas, terrestrial insect-eaters with spines, digging claws, and long sticky tongues. Although monotremes retain the ancestral trait of egg laying, they are not “primitive” relics; they are modern, specialized mammals that nourish their young with milk secreted onto the skin rather than through nipples.
Infraclass Marsupialia: Marsupials
Kingdom Animalia: Eumetazoa: Bilateria: Nephrozoa: Deuterostomia: Phylum Chordata: Subphylum Vertebrata: Superclass Gnathostomata: Clade Osteichthyes: Class Sarcopterygii: Clade Tetrapoda: Clade Amniota: Clade Synapsida: Class Mammalia: Clade Theria: Infraclass Marsupialia
Infraclass Marsupialia includes mammals such as kangaroos, wallabies, koalas, wombats, bandicoots, Tasmanian devils, and opossums. Marsupials are therian mammals, meaning they give birth to live young rather than laying eggs. However, unlike placental mammals, marsupials usually give birth after a short pregnancy to very underdeveloped young. The newborn then continues development while attached to a nipple, often inside a pouch called a marsupium. Not all marsupials have a large or obvious pouch, but extended development after birth is a key feature of the group. Marsupials are especially diverse in Australia and nearby regions, where they occupy many ecological roles as grazers, browsers, burrowers, climbers, predators, and insect eaters. They are also found in the Americas, especially in South America. The Virginia opossum is the only marsupial native to the United States and Canada, but many other opossum species live farther south. Marsupials show that live birth and parental care evolved in more than one successful form among mammals. Their reproductive strategy is different from that of placental mammals, but it is not inferior. It is one of several effective mammalian strategies for reproduction, development, and survival.
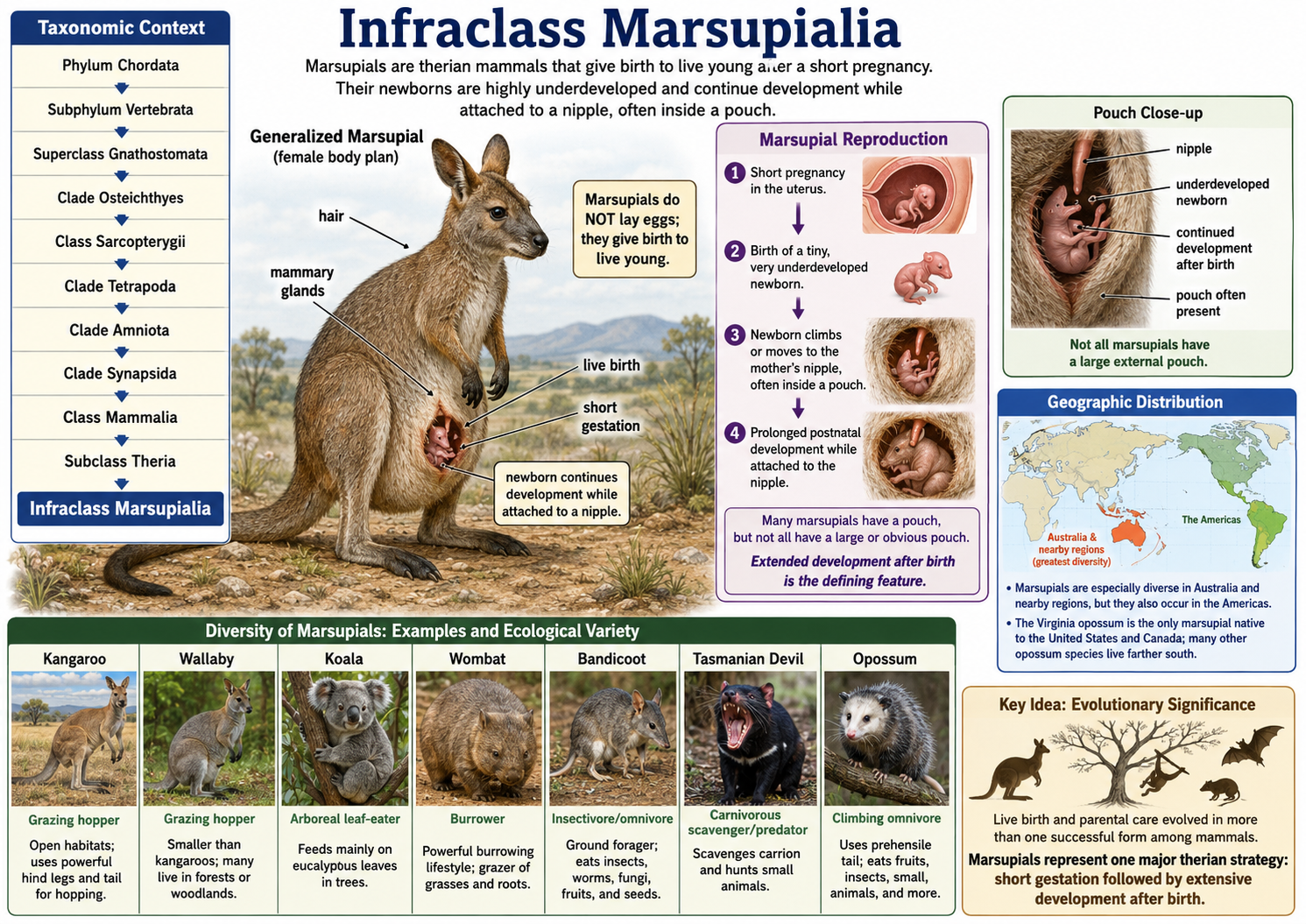
Figure 58. Infraclass Marsupialia. Mammals with extended postnatal development. Marsupials are live-bearing mammals that include kangaroos, wallabies, koalas, wombats, bandicoots, Tasmanian devils, and opossums. Their defining reproductive pattern is a short pregnancy followed by birth of a highly underdeveloped young that continues development while attached to a nipple, often within a pouch, although not all marsupials have a large external pouch. Marsupials are especially diverse in Australia and nearby regions but also occur in the Americas; the Virginia opossum is the only marsupial native to the United States and Canada. This group shows that mammals evolved more than one successful form of live birth and parental care.
Infraclass Eutheria: Placental Mammals
Kingdom Animalia: Eumetazoa: Bilateria: Nephrozoa: Deuterostomia: Phylum Chordata: Subphylum Vertebrata: Superclass Gnathostomata: Clade Osteichthyes: Class Sarcopterygii: Clade Tetrapoda: Clade Amniota: Clade Synapsida: Class Mammalia: Clade Theria: Infraclass Eutheria
Infraclass Eutheria includes the placental mammals and their close extinct relatives. Living placental mammals include humans, whales, dolphins, bats, rodents, rabbits, elephants, cats, dogs, horses, primates, and many others. Placental mammals are therian mammals, meaning they give birth to live young rather than laying eggs. They differ from marsupials because the embryo usually develops for a longer time inside the mother’s uterus, where it is supported by a complex placenta. The placenta is an organ that allows oxygen and nutrients to pass from the mother to the embryo and allows wastes to move from the embryo back to the mother. Because of this extended internal development, many placental mammals are born more developed than marsupial newborns. However, development at birth varies widely. A newborn horse can stand and walk soon after birth, while newborn mice, kittens, puppies, and humans require extensive parental care. Placental mammals are the most diverse living mammal group. They have adapted to many environments and ways of life. Bats evolved powered flight, whales and dolphins returned to aquatic life, moles became specialized for digging, and primates evolved grasping hands, forward-facing eyes, and complex social behavior. Humans are one branch of this much larger placental mammal radiation.
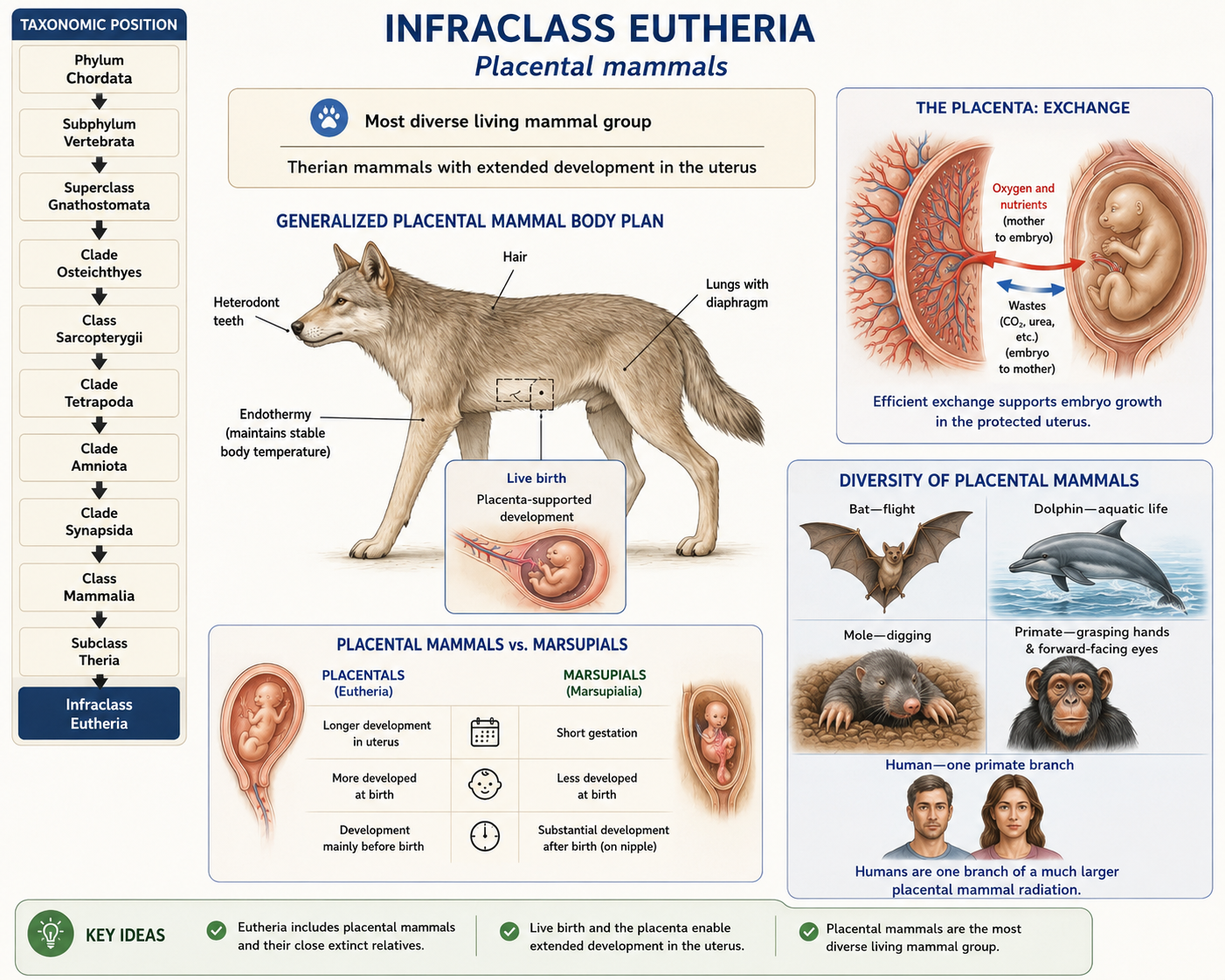
Figure 59. Infraclass Eutheria. Placental mammals. Eutherians include the placental mammals, the most diverse living mammal group, including humans, whales, bats, rodents, elephants, cats, dogs, and many others. Their defining reproductive feature is prolonged development of the embryo inside the mother’s uterus, where a placenta transfers oxygen and nutrients to the embryo and removes wastes. Compared with marsupials, placental mammals usually support more development before birth, so their young are often born in a more developed state. This lineage also shows extraordinary ecological diversity, including forms adapted for flight, aquatic life, digging, and complex primate behavior.